- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Image-Based Lignin Detection in Nematode-Induced Feeding Sites in Arabidopsis Roots

Published: Vol 15, Iss 9, May 5, 2025 DOI: 10.21769/BioProtoc.5301 Views: 2103
Reviewed by: Samik BhattacharyaPooja VermaShweta Panchal
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics









Abstract
Cyst and root-knot nematodes are sedentary biotrophic parasites that infect a wide range of plant species, causing significant annual yield and economic losses. Cyst nematodes (genera Heterodera and Globodera) induce specialized feeding structures called syncytia in host plant roots, while root-knot nematodes (Meloidogyne spp.) form galls containing feeding cells known as giant cells. This protocol describes the visualization of lignin in Arabidopsis roots infected by beet cyst nematode H. schachtii and root-knot nematode M. incognita using histochemical staining. We present two distinct approaches for lignin detection: direct staining of root segments containing syncytia and galls and histopathological detection in thin longitudinal sections of the feeding sites.
Key features
• First approach: Staining of intact roots visualizes lignin in nematode feeding sites and requires only simple specimen preparation and staining solution, with no sectioning needed.
• Second approach: Staining of longitudinal sections of feeding sites visualizes cell-specific lignin localization and requires moderate tissue preparation and sectioning.
• Both approaches enable specific detection and visualization of lignin in nematode-infected Arabidopsis tissues.
Keywords: Arabidopsis thalianaGraphical overview

Background
Plant-parasitic nematodes (PPNs) form a diverse group of microscopic roundworms and pose a serious threat to many crop plants, significantly impacting agricultural productivity [1,2]. Through their parasitic interactions, PPNs disrupt normal plant functions, leading to reduced nutrient and water uptake, stunted growth, and, in severe infestations, plant death [3].
PPNs secrete various enzymes and effectors that modify the host plant's cellular processes to support nematode development while suppressing the plant's immune responses [4]. Among the most damaging PPNs are sedentary root-knot (Meloidogyne spp.) and cyst nematodes (Heterodera spp. and Globodera spp.), as well as migratory lesion nematodes (Pratylenchus spp.) [5,6]. The endoparasitic nematodes induce sophisticated feeding sites in host roots, which serve as continuous nutrient sources for their development. While root-knot nematodes induce giant cells embedded in gall tissue, cyst nematodes establish so-called syncytia in plant roots.
The analyses of transcriptomes of syncytia and giant cells have shown that nematodes can suppress plant defense during the establishment of their feeding sites [7,8]. For instance, functional analysis of genes involved in pathways such as callose deposition, phytoalexin biosynthesis, reactive oxygen species (ROS) production, and antimicrobial peptide synthesis has revealed an overexpression of these genes in nematode-infected roots [9–12]. Similarly, genes related to lignin biosynthesis in the phenylpropanoid pathway have been studied in response to nematode infection, showing comparable results (Ali and Wieczorek, unpublished data). The phenylpropanoid pathway in plants leads to the biosynthesis of diverse secondary metabolites, including lignin, lignans, flavonoids, and phenolic compounds [13]. These compounds play essential roles in plant defense, pigmentation, structural integrity, rigidity, and the mechanical strength of cell walls [14]. Lignin, a key component of plant cell walls, is crucial for reinforcing structural integrity and serves as a protective barrier against invading pathogens [15]. Interestingly, DIR proteins, which are involved in lignin biosynthesis, do not function as enzymes. Instead, they guide the process by positioning monolignols in specific orientations, ensuring their proper coupling during polymerization [16]. This unique ability to control the spatial arrangement of lignin precursors facilitates the formation of a robust and intricate lignin network. Such a network not only strengthens the cell wall but also enhances plant defense by making it more difficult for pathogens to penetrate and infect the plant [17].
Disruption of lignin biosynthesis pathways has been shown to increase susceptibility to pathogens in Arabidopsis and other plants, highlighting the crucial role of these pathways in plant resistance [18]. Therefore, the present protocol has been developed to detect lignin in roots infected by the cyst nematode H. schachtii and the root-knot nematode M. incognita, which induce syncytia and galls, respectively. The staining method uses phloroglucinol-HCl, which reacts with the cinnamaldehyde end groups of lignin, producing a red-violet color. This method has not been widely used for staining lignin in nematode-induced feeding sites. Instead, many scientific publications analyze other cell wall polymers, such as cellulose and callose, using histochemical approaches or specific antibodies (e.g., [19,20]). However, Nakagami et al. (2020) used a 2% phloroglucinol solution to stain 3- and 5-day-old galls induced by M. incognita in Arabidopsis wild type and the mutant del1-1 [21]. The description of the method in this report is not very detailed. Additionally, the authors stained only whole root fragments containing the feeding sites and did not perform tissue sectioning to obtain a more detailed picture of lignin localization. Similarly, Sato et al. [22] used this method to stain entire 3-day-old galls induced by M. arenaria in Solanum torvum, demonstrating its applicability to Solanum species. However, the description of the staining protocol in this study is not detailed, and the authors refer to a histochemistry book by Jensen (1962) [23], which is not easily accessible. Hence, this protocol provides, for the first time, a detailed description of lignin staining in both whole nematode-feeding sites and sectioned feeding sites induced in Arabidopsis, allowing for a more precise localization of lignin deposition. Its comprehensive methodology enhances reproducibility and enables a clearer understanding of cell wall modifications in response to nematode infection.
Materials and reagents
Biological materials
1. Arabidopsis thaliana (Col-0)
2. Beet root cyst nematode Heterodera schachtii, originally from the Institute of Phytopathology (group of Prof. Florian Grundler), Christian-Albrecht University, Kiel, Germany
3. Root-knot nematode Meloidogyne incognita, originally from the Institute of Phytopathology (group of Prof. Florian Grundler), Christian-Albrecht University, Kiel, Germany
Reagents
1. Type 2 distilled water (dH2O)
2. Ethanol
3. Sodium hypochlorite (Carl Roth, catalog number: 9062.3)
4. Phloroglucinol (Merck, catalog number: 7069)
5. Formaldehyde (Carl Roth, catalog number: 4980.1)
6. Low-melting agarose (Seaplague Agarose) (Duchefa, catalog number: S1202.0100)
7. Saccharose (table sugar) (AGRANA Zucker GmbH, Wien, Austria)
8. Daishin agar (Duchefa, catalog number: D004)
9. Gelrite (Duchefa, catalog number: G1101)
10. Gamborg B5 vitamin mixture (Duchefa, catalog number: G0415)
11. MES (Carl Roth, catalog number: 4259.3)
12. 37% HCl (Chem-Lab, CAS: 7647-01-0)
13. KNO3 (Sigma-Aldrich, catalog number: 31263)
14. MgSO4·7H2O (Carl Roth, catalog number: 8283.1)
15. Ca(NO3)·4H2O (Carl Roth, catalog number: P740.3)
16. KH2PO4 (Carl Roth, catalog number: 3904.3)
17. FeNaEDTA (Carl Roth, catalog number: 6498.1)
18. HBO3 (Merck, catalog number: B0394)
19. MnCl2 (Carl Roth, catalog number: T881.3)
20. CuSO4 (Sigma-Aldrich, catalog number: C8027)
21. ZnSO4 (Carl Roth, catalog number: T884.1)
22. CoCl2 (Sigma-Aldrich, catalog number: C866.1)
23. H2MoO4 (Carl Roth, catalog number: 2722.1)
24. NaCl (Carl Roth, catalog number: 9265.1)
25. NaH2PO4 (Carl Roth, catalog number: T879.2)
26. HgCl2 (Carl Roth, catalog number: 7904.1)
Solutions
1. B5 solid medium (see Recipes)
2. 0.2× Knop solid medium (see Recipes)
3. 0.2× Knop stock solution I (see Recipes)
4. 0.2× Knop stock solution II (see Recipes)
5. 0.2× Knop stock solution III (see Recipes)
6. 0.2× Knop stock solution IV (see Recipes)
7. 0.2× Knop stock solution V (see Recipes)
8. 10× phosphate-buffered saline (PBS) (see Recipes)
9. Fixation solution (see Recipes)
10. 10× washing buffer (see Recipes)
11. 2% phloroglucinol solution (see Recipes)
Recipes
1. B5 solid medium
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| Saccharose | 20 g/L | 20 g |
| Daishin agar | 8 g/L | 8 g |
| Gamborg B5 vitamin mixture | n/a | 1 mL |
| MES | 0.5 g/L | 0.5 g |
| Total | n/a | 1,000 mL |
Adjust the pH to 5.7 using 1 M KOH solution. If the pH exceeds the desired level due to the inadvertent addition of excess KOH, it is permissible to readjust the pH by carefully adding HCl or another suitable acid. Autoclave the B5 medium and pour approximately 50 mL into Petri dishes with cams (150 × 20 mm) inside a sterile laminar flow hood on the same day.
2. 0.2× Knop solid medium
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| Saccharose | 20 g/L | 20 g |
| Daishin Agar | 8 g/L | 8 g |
| Gamborg B5 vitamin mixture | n/a | 1 mL |
| Stock solution I | n/a | 2 mL |
| Stock solution II | n/a | 2 mL |
| Stock solution III | n/a | 0.4 mL |
| Stock solution IV | n/a | 0.2 mL |
| Stock solution V | n/a | 1 mL |
| Total | n/a | 1,000 mL |
Adjust the pH to 6.4 using 1 M KOH solution. If the pH exceeds the desired level due to the inadvertent addition of excess KOH, it is permissible to readjust the pH by carefully adding HCl or another suitable acid. Autoclave the 0.2× Knop medium and pour approximately 20 mL into Petri dishes with cams (92 × 16 mm) inside a sterile laminar flow hood on the same day.
3. 0.2× Knop stock solution I
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| KNO3 | 600 mM | 60.66 g |
| MgSO4·7H2O | 40 mM | 9.855 g |
| Total | n/a | 1,000 mL |
4. 0.2× Knop stock solution II
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| Ca(NO3)2·4H2O | 254 mM | 60 g |
| Total | n/a | 1,000 mL |
5. 0.2× Knop stock solution III
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| KH2PO4 | 100 mM | 13.61 g |
| Total | n/a | 1,000 mL |
6. 0.2× Knop stock solution IV
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| FeNaEDTA | 8.7 mM | 3.67 g |
| Total | n/a | 1,000 mL |
7. 0.2× Knop stock solution V
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| H3BO3 | 23.1 mM | 1.43 g |
| MnCl2 | 7.2 mM | 0.905 g |
| CuSO4·5H2O | 0.15 mM | 0.0365 g |
| ZnSO4·7H2O | 0.63 mM | 0.18 g |
| CoCl2·6H2O | 0.6 mM | 0.015 g |
| H2MoO4 | 0.16 mM | 0.026 g |
| NaCl | 17 mM | 1 g |
| Total | n/a | 1,000 mL |
8. 10× phosphate-buffered saline (PBS)
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| NaH2PO4 | 10 mM | 1.41 g |
| NaCl | 130 mM | 7.6 g |
| dH2O | n/a | n/a |
| Total (optional) | n/a | 1,000 mL |
Adjust the pH to 7.5 using 1 M KOH solution.
9. Fixation solution
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| Ethanol | 63% | 6.3 mL |
| Formaldehyde in PBS | 5% | 0.5 mL |
| dH2O | n/a | 3.2 mL |
| Total (optional) | n/a | 10 mL |
Adjust the pH to 7.2 using 1 M KOH solution.
10. Washing buffer
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| Ethanol | 63% | 630 mL |
| PBS | n/a | 370 mL |
| Total (optional) | n/a | 1,000 mL |
11. 2% phloroglucinol solution
| Reagent | Final concentration | Quantity or Volume |
|---|---|---|
| Ethanol | 100% | 10 mL |
| Phloroglucinol | 2% | 0.2 g |
| Total (optional) | n/a | 10 mL |
Subsequently, mix two parts of 2% phloroglucinol solution with one part of 37% HCl.
Laboratory supplies
1. Parafilm (Bemis, model: PM 996)
2. Petri dishes with cams (92 × 16 mm) (Sarstedt, catalog number: 82.1473)
3. Petri dishes with cams (150 × 20 mm) (Sarstedt, catalog number: 82.1184.500)
4. Petri dishes without cams (54.65 × 14.7 mm) (Sarstedt, catalog number: 82.9923.426)
5. 12-well plates (Sarstedt, catalog number: 83.3921)
6. Quick-drying glue (e.g., Superglue, Loctite, Germany)
Equipment
1. Vibratome (Leica, model: VT100S) with stereomicroscope (in this protocol, Olympus, model: SZ51)
2. Heating plate
3. Laminar flow hood
4. Microscope with integrated camera (in this protocol, Olympus, model: BX53 with U-CMAD3 camera)
Software and datasets
1. Olympus cellSens Dimension 3.2 (Olympus, Tokyo, Japan)
Procedure
Note: This protocol can be adapted for other host plants infected with different plant-parasitic nematodes. However, specific adjustments may be required, such as modifications to the fixation and staining duration, as well as settings for section preparation using the vibratome.
A. H. schachtii stock culture on mustard and inoculation of Arabidopsis plants
For more details on H. schachtii stock culturing on mustard and the inoculation of Arabidopsis plants, please see the protocol by Bohlmann and Wieczorek [24]. Briefly, the stock culture was maintained on mustard plants grown on 0.2× Knop medium in Petri dishes with cams (150 × 20 mm). Twelve-day-old Arabidopsis plants grown in Petri dishes were inoculated with freshly hatched juveniles of H. schachtii.
B. M. incognita stock culture on cucumber and inoculation of Arabidopsis plants
1. In vitro stock culturing of M. incognita is done on cucumber as follows:
a. Surface-sterilize the cucumber seeds with 6% sodium hypochlorite solution for 1 h, followed by washing in sterile dH2O for 1 h. Repeat these steps three times. Subsequently, wash the seeds for 1 min in 70% ethanol.
b. Wash the seeds three to four times with autoclaved sterile dH2O to remove the traces of sodium hypochlorite and leave them in the laminar flow hood until the seeds are completely dry.
c. Place several sterilized seeds into a germination plastic box with a sterile wet tissue at the bottom. Let the seeds germinate for 10–14 days at room temperature in the dark.
d. Transfer three cucumber seedlings from the germination box into a 150 × 20 mm Petri dish containing 0.2× Knop medium supplemented with vitamin B5. Trim the shoots of the seedlings to avoid damaging the B5 medium (see Recipes). Incubate the plates in the dark at room temperature for two weeks.
e. Prior to inoculation, pick the egg masses from older stock plates and put them into the hatching funnel as described for H. schachtii in the protocol by Bohlmann and Wieczorek [19]. Use sterile dH2O instead of 3 mM ZnCl2 to stimulate hatching.
f. After 4–5 days, collect the freshly hatched second-stage juveniles, sterilize them with 0.01% HgCl2, and wash them three times with sterile dH2O. Subsequently, resuspend the larvae in the 0.7% (w/v) gelrite.
g. Use approximately 500 juveniles for three cucumber roots and subsequently cultivate the plates in the dark cabinet at room temperature until further use without sub-culturing.
h. Newly produced egg masses can be harvested after 2–3 months. Already hatched second-stage juveniles can be washed off the medium surface with sterile dH2O.
2. Inoculation of Arabidopsis with M. incognita
a. Prepare 50 mL of sterilization solution containing 25 mL of 70% ethanol, 5 mL of 12% sodium hypochlorite, and 20 mL of sterile dH2O.
b. Surface-sterilize the Arabidopsis seeds with a freshly prepared sterilization solution for 3–5 min followed by 3× washing steps with sterile dH2O. Subsequently, wash the seeds in 70% ethanol for 1 min.
c. Pour approximately 20 mL of 0.2× Knop medium in 92 × 16 mm Petri dishes.
d. Place 10 sterilized seeds on 0.2× Knop medium per dish, arranging them in two lines of five seeds in the upper half of the dish. Incubate the dishes in a growth chamber set to 22–25 °C with a photoperiod of 16/8 h light/dark. For more details, see the protocol by Bohlmann and Wieczorek [24].
e. Tilt the dishes at a 45–60° angle after germination to allow the roots to grow vertically (Figure 1A).
f. Collect the egg masses from the stock culture and place them in a small sterile beaker with sterile dH2O. Incubate them in the dark for 3–4 days at room temperature to induce hatching of second-stage juveniles.
g. Resuspend the sterile larva in 0.7% (w/v) gelrite and mix thoroughly to obtain a homogenous solution.
h. Count the number of second-stage juveniles per drop (e.g., 10 or 20 μL). The optimal number of juveniles ranges between 15–20 per drop.
i. Distribute a few drops of gelrite next to the root tips of the 12-day-old plants (Figure 1B) containing a defined number of second-stage juveniles, resulting in an inoculation of approximately 50–60 juveniles per plant.
j. Place the Petri dishes back in the growth chamber set to 22–25 °C with a photoperiod of 16/8 h light/dark.
k. Collect control and infected root segments at the desired time point (depending on the research question; here, 9 and 14 days after nematode inoculation) and proceed with fixation and lignin staining.
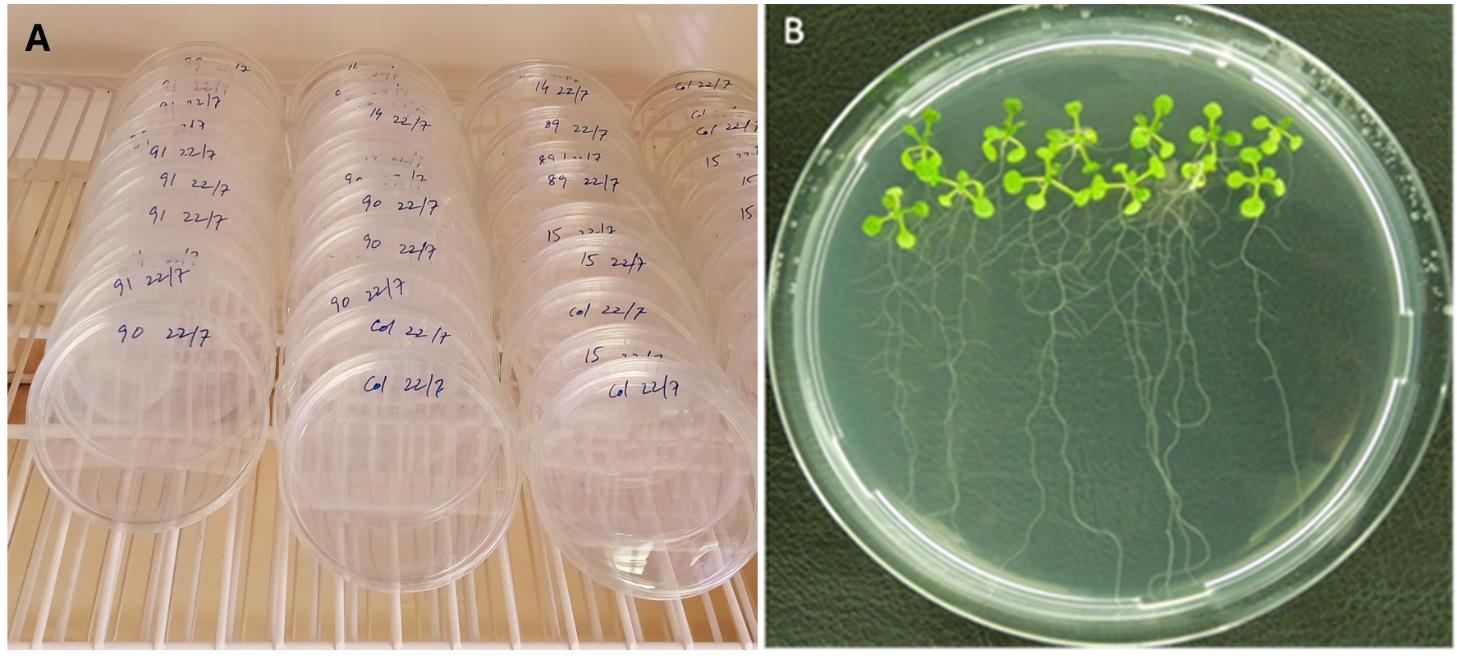
Figure 1. Cultivation of Arabidopsis plants on 0.2× Knop medium. (A) Tilting of dishes at a 45–60° angle. (B) Petri dish containing 10 plants, 12 days after germination, ready for inoculation with nematodes.
C. Fixation and washing
1. Using a scalpel and forceps, carefully cut out control and infected Arabidopsis root segments that contain syncytia and galls. Place the segments into Eppendorf tubes (1.5 mL or 2 mL) filled with pre-chilled fixation solution (refer to Recipes) (Figure 2A). In this protocol, root segments were collected at 9 and 14 days post-nematode inoculation. Vacuum infiltrate the fixation solution on ice for 15–20 min to ensure the proper penetration of the fixative into the tissues (Figure 2B).
2. Incubate samples for 42 h or longer at 4 °C on a shaker with gentle shaking (Figure 2C).
3. After fixation, wash the samples thrice with washing buffer (see Recipes) for 10 min, followed by one wash in 1% PBS for 10 min (Figure 2D).

Figure 2. Fixation and washing of uninfected and infected Arabidopsis root samples containing syncytia and galls induced by H. schachtii and M. incognita, respectively. (A) Fixation of the root segments in cold fixation solution. (B) Vacuum infiltration of the fixative. (C) Shaking of samples at 4 °C for 40 h. (D) Washing with PBS buffer.
D. Lignin staining protocol for intact nematode-infected Arabidopsis roots
1. After the last washing step, remove the root samples from 1% PBS using forceps and briefly transfer them to a paper towel for a few seconds to remove any remaining PBS solution.
2. Place 5–10 segments per well in a 12-well plate containing 2% phloroglucinol solution (see Recipes) and vacuum infiltrate for 10–15 min for detection of lignin.
3. Subsequently, wash the root segments 2–3 times with sterile dH2O.
4. Photograph the stained roots, syncytia, and galls at the required magnification (here, 40×), e.g., using an Olympus BX53 microscope with an integrated camera U-CMAD3.
E. Lignin staining protocol for longitudinal sections of nematode-induced syncytia and galls in Arabidopsis roots
For more details on embedding and sectioning using a vibratome, see the protocol by Wieczorek [25].
1. Embedding
a. Prepare 5% (w/v) low-melting agarose in PBS buffer (see Recipes; Figure 3A).
b. Place a heating plate in the laminar flow hood and set it to 35–40 °C (Figure 3B).
c. Pour low-melting agarose into a small petri dish (54.65 × 14.7 mm) placed on a prewarmed heating plate.
d. Grab the root segments using forceps (Figure 3C), carefully swab them with a paper towel to remove excess PBS (Figure 3D), and dip the root segments into warm agarose parallel to the bottom of the Petri dish, approximately halfway into the agarose (Figure 3E).
e. Use approximately 10 root segments per dish (Figure 3F). Throughout the procedure, keep the dish on a heating plate to maintain the agarose at the appropriate temperature and prevent solidification.
f. After embedding all root segments, remove the Petri dish from the heating plate and allow the agarose to cool and solidify inside the laminar flow hood.
g. Subsequently, seal the Petri dish with parafilm and store it at 4 °C for several weeks before sectioning.

Figure 3. Embedding of control and nematode-infected Arabidopsis root segments. (A) Flask with low-melting agarose on a heating plate. (B) A Petri dish on a heating plate. (C) Picking up root segments from 1% PBS buffer. (D) Swabbing of root segments with a paper towel. (E) Embedding the root segments in low-melting agarose. (F) Approximately 10 root segments embedded in one Petri dish.
2. Preparation of longitudinal sections of nematode-induced feeding sites (using syncytia induced by H. schachtii as an example) with a vibratome (Figure 4A)
a. Cut out small agarose blocks (approximately 10 mm high × 6–8 mm wide) containing a single root segment with a scalpel.
b. Trim the sides of the agarose block under the microscope, leaving some agarose around the root segments.
c. Glue the agarose block with the root segment onto the round vibratome plate with a drop of quick-drying glue.
d. To produce longitudinal sections, the root segment within the agarose block must be placed parallel to the bottom of the vibratome plate.
e. Cut 20–25 μm thick tissue sections in the vibratome tray filled with dH2O (Figure 4B; in this protocol, we used a cutting speed at level 2–3 and frequency at level 5–6). Keep a close eye on the sections using the microscope while cutting (here, Olympus SZ51 stereomicroscope with a 10× objective and 1× auxiliary lens) and pick the sections one by one using feather forceps.
f. Collect the sections in a watch glass or 12-well plate filled with dH2O.
g. Transfer a longitudinal section of syncytia into a drop of dH2O placed on a slide (Figure 4C).
h. Place the slide on a heating plate set to 30–40 °C to allow the dH2O to slowly evaporate. To flatten the longitudinal sections, use a cotton bud dipped in acetone and move it slowly above the water drop without touching it (Figure 4D).
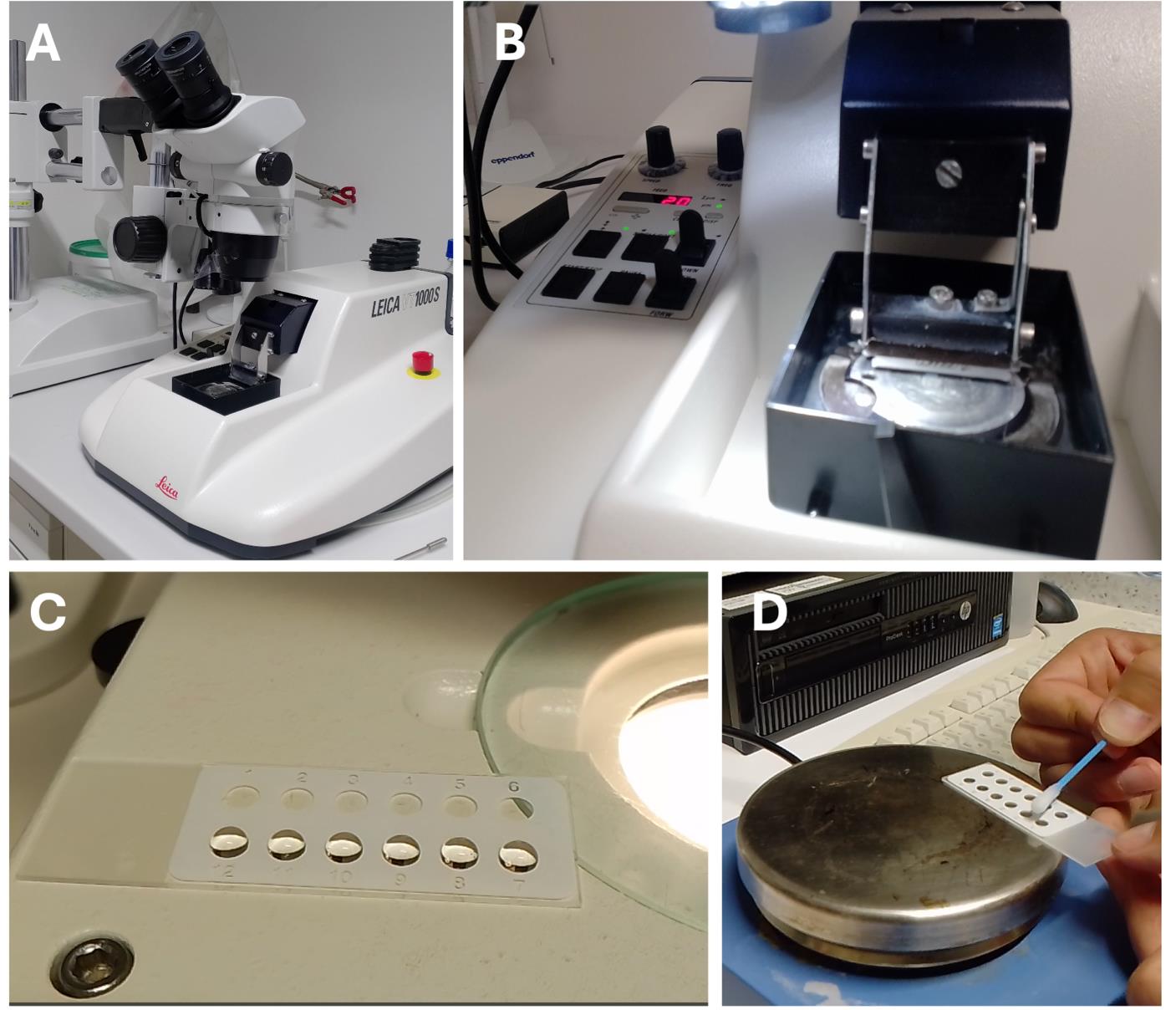
Figure 4. Preparation of longitudinal sections of syncytia induced by H. schachtii in Arabidopsis roots . (A) Vibratome with a stereomicroscope. (B) Preparation of longitudinal tissue sections. (C) Collecting longitudinal sections on a glass slide. (D) Flattening longitudinal sections using a cotton bud dipped in acetone.
3. Lignin staining of longitudinal sections
a. Add one drop of phloroglucinol solution to each longitudinal section and incubate for 10–20 min.
b. Subsequently, carefully wash the longitudinal sections with dH2O, place a coverslip over them, and photograph them under a microscope equipped with an integrated camera, e.g., Olympus BX53 microscope with an integrated camera U-CMAD3 connected to a computer with the CellSens software.
Validation of protocol
The first part of this protocol was applied in our laboratory to gain new insights into the biosynthesis and localization of lignin in intact, un-sectioned 9- and 14-day-old feeding sites induced by cyst- (Figure 5A, B) and root-knot nematodes in Arabidopsis roots (Figure 5C, D). The corresponding stained uninfected control roots are depicted in Figure 5C, F.
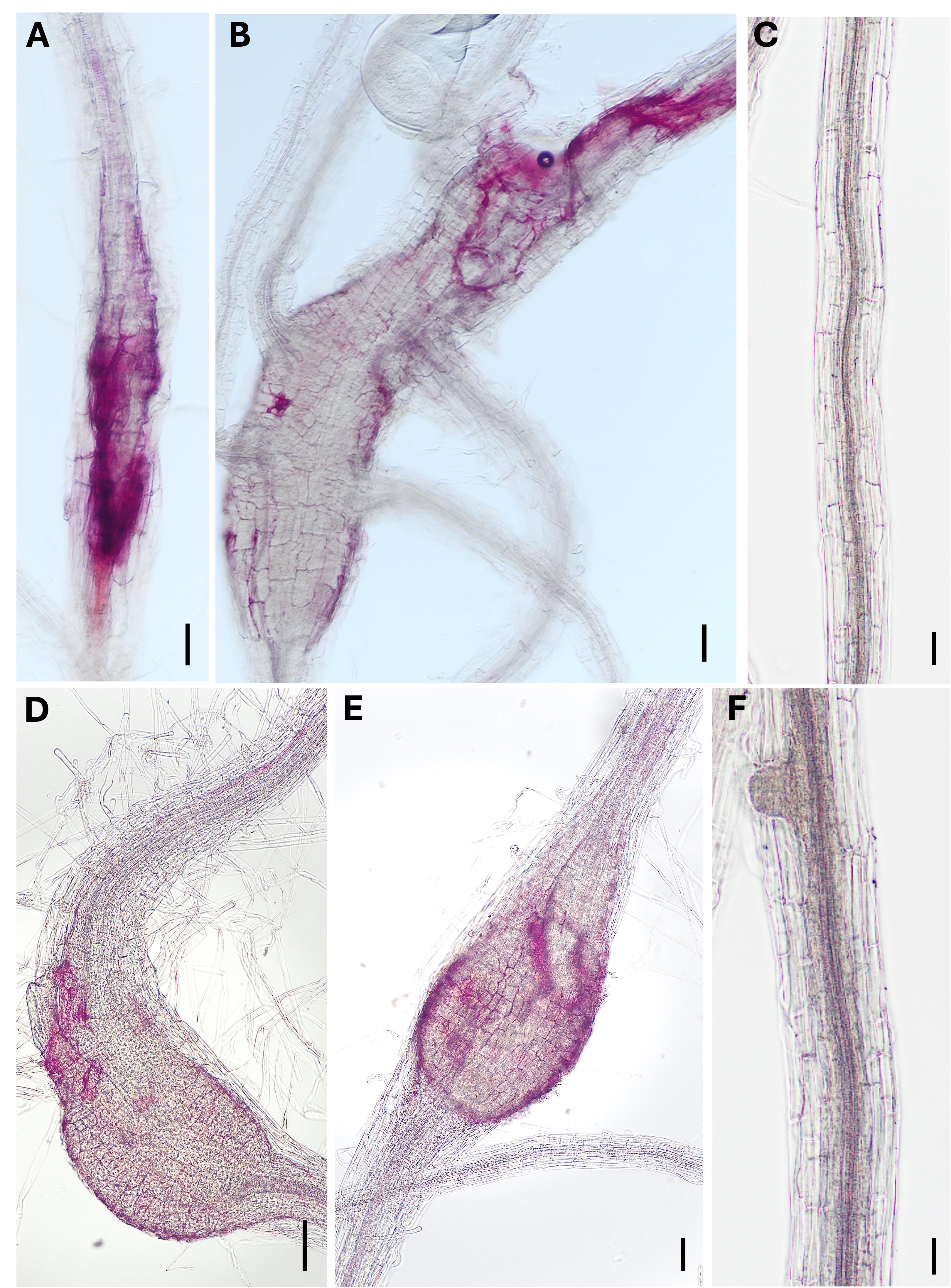
Figure 5. Lignin staining in nematode-infected Arabidopsis roots. Root segments containing 9- and 14-day-old syncytia (A and B) and galls (D and E) induced by H. schachtii and M. incognita, respectively, as well as uninfected 9- (C) and 14-day-old (F) control roots. Scale bars = 100 μm.
The second part of this protocol was applied in our laboratory to gain further insights into the specific localization of lignin in longitudinal sections of 9- and 14-day-old feeding sites induced by cyst- and root-knot nematodes in Arabidopsis roots. Figure 6A, B shows examples of stained syncytia induced by H. schachtii, and Figure 6C shows a stained segment section of uninfected roots.
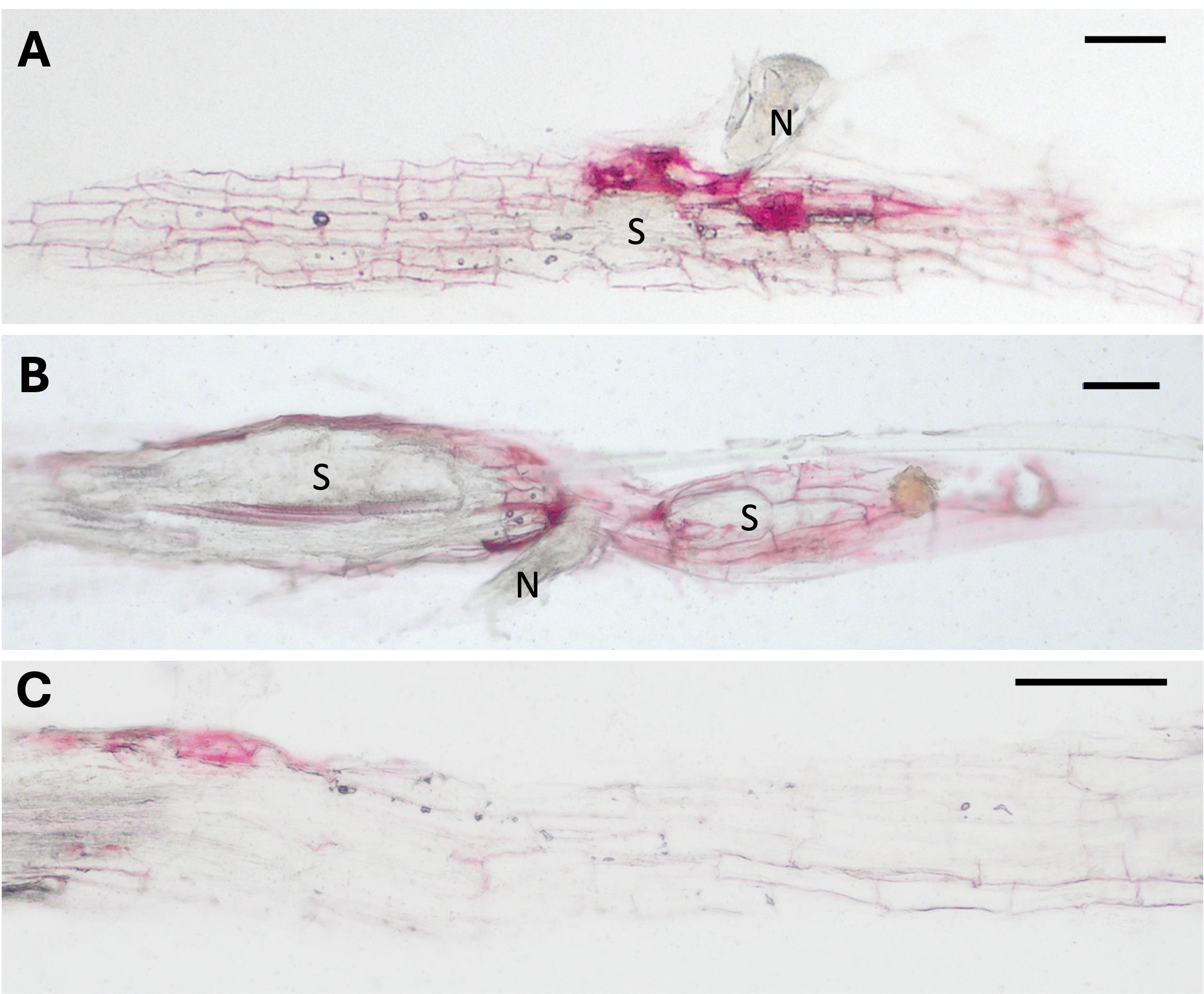
Figure 6. Lignin staining in longitudinal syncytia sections in Arabidopsis roots. (A) 9- and (B) 14-day-old feeding sites induced by H. schachtii and (C) the uninfected control root. N: nematode; S: syncytium. Scale bars = 100 μm.
Other parts of this protocol, such as cyst and root-knot nematode stock culture, inoculation of Arabidopsis plants with H. schachtii and M. incognita, and preparation of thin sections of nematode feeding sites, have been validated in the following research articles:
• Bohlmann et al. [24]. Infection Assay of Cyst Nematodes on Arabidopsis Roots. Bio-protoc. 5(18): e1596. https://doi.org/10.21769/BioProtoc.1596
• Wieczorek [25]. Detection and Visualization of Specific Gene Transcripts by in situ RT-PCR in Nematode-Infected Arabidopsis Root Tissue. Bio-protoc. 5(18): e1597. https://doi.org/10.21769/BioProtoc.1597
• Wieczorek et al. [20]. A distinct role of pectate lyases in the formation of feeding structures induced by cyst and root-knot nematodes. Mol. Plant Microbe Interact. 27: 901–912. https://doi.org/10.1094/MPMI-01-14-0005-R
• Opitz et al. [26]. The other side of the coin: systemic effects of Serendipita indica root colonization on development of sedentary plant–parasitic nematodes in Arabidopsis thaliana. Planta. 259: 121. https://doi.org/10.1007/s00425-024-04402-5
Acknowledgments
Muhammad Amjad Ali was supported by Ernst Mach grant (EZA) number MPC-2023-06337 from OeAD-GmbH, Vienna, Austria. We gratefully acknowledge Tina Austerlitz for her outstanding assistance in the laboratory.
Competing interests
The authors declare no conflicts of interest.
References
- Jones, J. T., Haegeman, A., Danchin, E. G. J., Gaur, H. S., Helder, J., Jones, M. G. K., Kikuchi, T., Manzanilla-López, R., Palomares-Rius, J. E., Wesemael, W. M. L. et al. (2013). Top 10 plant-parasitic nematodes in molecular plant pathology. Mol Plant Pathol. 14(9): 946–961. https://doi.org/10.1111/mpp.12057
- Ali, M. A., Azeem, F., Abbas, A., Joyia, F. A., Li, H., Dababat, A. A. (2017a). Transgenic strategies for enhancement of nematode resistance in plants. Front Plant Sci. 8: 750. https://doi.org/10.3389/fpls.2017.00750
- Davis, E. L., Hussey, R. S. and Baum, T. J. (2004). Getting to the roots of parasitism by nematodes. Trends Parasitol. 20(3): 134–141. https://doi.org/10.1016/j.pt.2004.01.008
- Zahoor, A., Niaz, Z., Tahir-ul-Qamar, M. and Hawamda, A. I. M. (2023). Computational analysis of 16D10 effector peptides from root-knot nematodes and their interaction with Mi resistance (R) protein through molecular docking. J Integr Plant Biotech. 1(1): 01–08. https://doi.org/10.55627/pbiotech.001.01.0665
- Moens, M., Perry, R. N. and Starr, J. L. (2009). Meloidogyne species: a diverse group of novel and important plant parasites. In Perry, R. N., Moens, M. and Starr J. L. (Eds.). Root-knot Nematodes. Wallingford, UK: CABI Publishing. 1–17.
- Nicol, J. M., Turner, S. J., Coyne, D. L., den Nijs, L., Hockland, S. and Maafi, Z. T. (2011). Current nematode threats to world agriculture. In Jones, J. T., Gheysen, G. and Fenoll, C. (Eds.). Genomics and Molecular Genetics of Plant–Nematode Interactions. Springer. 21–44.
- Ali, M. A., Abbas, A., Azeem, F., Javed, N. and Bohlmann, H. (2015). Plant-nematode interactions: From genomics to metabolomics. Int J Agric Biol. 17(6): 1071–1082. https://doi.org/10.17957/IJAB/15.0001
- Ali, M. A., Azeem, F., Li, H. and Bohlmann, H. (2017b). Smart parasitic worms use multifaceted strategies to parasitize plants. Front Plant Sci. 8: 1699. https://doi.org/10.3389/fpls.2017.01699
- Ali, M. A., Abbas, A., Kreil, D. P. and Bohlmann, H. (2013). Overexpression of the transcription factor RAP2.6 leads to enhanced callose deposition in syncytia and enhanced resistance against the beet cyst nematode Heterodera schachtii in Arabidopsis roots. BMC Plant Biol. 13: 47. https://doi.org/10.1186/1471-2229-13-47
- Ali, M. A., Wieczorek, K., Kreil, D. P. and Bohlmann, H. (2014). The beet cyst nematode Heterodera schachtii modulates the expression of WRKY transcription factors in syncytia to favour its development in Arabidopsis roots. PLoS One. 9(7): e102360. https://doi.org/10.1371/journal.pone.0102360
- Ali, M. A., Abbas, A., Zahoor, A., Azeem, F. and Bohlmann, H. (2020). The Arabidopsis GPI anchored LTPg5 encoded by At3g22600 is involved in plant defense against the beet cyst nematode Heterodera schachtii. Int J Mol Sci. 21(5): 1774. https://doi.org/10.3390/ijms21051774
- Hawamda, A. I. M., Zahoor, A., Abbas, A., Ali, M. A. and Bohlmann, H. (2020). The Arabidopsis RbohB encoded by At1g09090 is important for resistance against nematodes. Int J Mol Sci. 21(15): 5556. https://doi.org/10.3390/ijms21155556
- Dixon, R. A. and Paiva, N. L. (1995). Stress-induced phenylpropanoid metabolism. Plant Cell. 7(7): 1085–1097. https://doi.org/10.1105/tpc.7.7.1085
- Hatfield, R. D., Grabber, J. H., Ralph, J. and Vermerris, W. (2016). Lignin formation in plant cell walls: Key roles for p-coumaroylation and feruloylation in cell wall assembly and resistance. Plant Physiol. 140(4): 948–956. https://doi.org/10.1104/pp.105.072579
- Vanholme, R., Demedts, B., Morreel, K., Ralph, J. and Boerjan, W. (2010). Lignin Biosynthesis and Structure. Plant Physiol. 153(3): 895–905. https://doi.org/10.1104/pp.110.155119
- Davin, L. B. and Lewis, N. G. (2005). Dirigent phenoxy radical coupling: Advances and challenges. Curr. Opin. Biotech., 16(4): 398–406. https://doi.org/10.1016/j.copbio.2005.06.010
- Miedes, E., Vanholme, R., Boerjan, W. and Molina, A. (2014). The role of the secondary cell wall in plant resistance to pathogens. Front Plant Sci. 5: 358. https://doi.org/10.3389/fpls.2014.00358
- Gallego‐Giraldo, L., Posé, S., Pattathil, S., Peralta, A. G., Hahn, M. G., Ayre, B. G., Sunuwar, J., Hernandez, J., Patel, M., Shah, J., et al. (2018). Elicitors and defense gene induction in plants with altered lignin compositions. New Phytol. 219(4): 1235–1251. https://doi.org/10.1111/nph.15258
- Meidani, C., Ntalli, N. G., Giannoutsou, E. and Adamakis, I.-D. S. (2019). Cell Wall Modifications in Giant Cells Induced by the Plant Parasitic Nematode Meloidogyne incognita in Wild-Type (Col-0) and the fra2 Arabidopsis thaliana Katanin Mutant. Int J Mol Sci. 20: 5465. https://doi.org/10.3390/ijms20215465
- Wieczorek, K., Elashry, A., Quentin, M., Grundler, F. M. W., Favery, B., Seifert, G. J. and Bohlmann, H. (2014). A distinct role of pectate lyases in the formation of feeding structures induced by cyst and root-knot nematodes. Mol Plant Microbe Interact. 27: 901–912. https://doi.org/10.1094/MPMI-01-14-0005-R
- Nakagami, S., Saeki, K., Toda, K., Ishida, T. and Sawa, S. (2020). The atypical E2F transcription factor DEL1 modulates growth–defense tradeoffs of host plants during root-knot nematode infection. Sci Rep. 10(1): 8836. https://doi.org/10.1038/s41598-020-65733-3
- Sato, K., Uehara, T., Holbein, J., Sasaki-Sekimoto, Y., Gan, P., Bino, T., Yamaguchi, K., Ichihashi, Y., Maki, N., Shigenobu, S., et al. (2021). Transcriptomic analysis of resistant and susceptible responses in a new model root-knot nematode infection system using Solanum torvum and Meloidogyne arenaria. Front Plant Sci. 12: 980. https://doi.org/10.3389/fpls.2021.680151
- Jensen, W. A. (1962). Botanical Histochemistry. London: W.H. Freeman & Co.
- Bohlmann, H. and Wieczorek, K (2015). Infection Assay of Cyst Nematodes on Arabidopsis Roots. Bio-protoc. 5(18): e1596. https://doi.org/10.21769/BioProtoc.1596
- Wieczorek, K. (2015). Detection and Visualization of Specific Gene Transcripts by in situ RT-PCR in Nematode-Infected Arabidopsis Root Tissue. Bio-protoc. 5(18): e1597. https://doi.org/10.21769/BioProtoc.1597
- Opitz, M., Díaz-Manzano, F. E., Ruiz-Ferrer, V., Daneshkhah, R., Lorenz, C., Ludwig, R., Escobar, C., Steinkellner, S., Wieczorek, K. (2024). The other side of the coin: systemic effects of Serendipita indica root colonization on development of sedentary plant–parasitic nematodes in Arabidopsis thaliana. Planta. 259: 121. https://doi.org/10.1007/s00425-024-04402-5
Article Information
Publication history
Received: Feb 17, 2025
Accepted: Apr 7, 2025
Available online: Apr 20, 2025
Published: May 5, 2025
Copyright
© 2025 The Author(s); This is an open access article under the CC BY-NC license (https://creativecommons.org/licenses/by-nc/4.0/).
How to cite
Ali, M. A. and Wieczorek, K. (2025). Image-Based Lignin Detection in Nematode-Induced Feeding Sites in Arabidopsis Roots. Bio-protocol 15(9): e5301. DOI: 10.21769/BioProtoc.5301.
Category
Plant Science > Plant immunity > Plant-insect interaction
Plant Science > Plant physiology > Nodulation
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.