- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Measurement of Reactive Oxygen and Nitrogen Species in Living Cells Using the Probe 2',7'-Dichlorodihydrofluorescein
Published: Vol 11, Iss 24, Dec 20, 2021 DOI: 10.21769/BioProtoc.4279 Views: 4129
Reviewed by: ASWAD KHADILKARManasi K. MayekarArvind Panday
Original research article
The authors used this protocol in:
Jan 2019
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols
Abstract
Reactive oxygen species and reactive nitrogen species (RONS) are involved in programmed cell death in the context of numerous degenerative and chronic diseases. In particular, the ability of cells to maintain redox homeostasis is necessary for an adaptive cellular response to adverse conditions that can cause damage to proteins and DNA, resulting in apoptosis and genetic mutations. Here, we focus on the 2',7'-dichlorodihydrofluorescein diacetate (DCFH2-DA) assay to detect RONS. Although this fluorescence-based assay is widely utilized due to its high sensitivity to detect changes in cellular redox status that allow measuring alterations in RONS over time, its validity has been a matter of controversy. If correctly carried out, its limitations are understood and results are correctly interpreted, the DCFH2-DA assay is a valuable tool for cell-based studies.
Background
Reactive oxygen and nitrogen species (RONS) is a shared term for oxygen-based radicals and/or chemically reactive oxidants including nitric oxide (•NO), superoxide (O2•−), hydrogen peroxide (H2O2), peroxynitrite (ONOO−), hypochlorous acid (HOCl), and hydroxyl radical (•OH), among others (Halliwell, 2012). RONS, produced mainly from nicotinamide adenine dinucleotide phosphate oxidase and nitric oxide synthase, play an essential role in signaling cascades regulating cellular processes such as proliferation, differentiation, motility, and programmed cell death (Dröge, 2002). Of note, in view of a cellular vulnerability to oxidants or activators of oxidative metabolism that can induce cell death, an usually tightly regulated steady-state control over the production-detoxification of RONS protects cells from their harmful effects (Harris and Brugge, 2015). This has stimulated increased investigations and lead to tools that are aimed at a better understanding of the biochemistry and physiology of redox reactions of the RONS. However, these tools are not able to assess all facets of redox mechanisms, specifically in living cells and organisms, since detection and quantification of short-lived RONS requires practices that respond to them very rapidly, as opposed to classical antioxidants that finally generate stable products (Dikalov and Harrison, 2014; Brandes et al., 2018). While RONS and redox biology are associated with complex systems, assays that are used for the measurement of RONS involved in biological systems should be approached with caution.
Fluorescent probes produce stable products upon reactions with RONS and can offer high sensitivity detection (Hempel et al., 1999). Redox signaling mechanisms can be explored using 2',7'- dichlorodihydrofluorescein diacetate (DCFH2-DA); this cell-permeable ester is hydrolyzed inside the cell to 2',7'-dichlorodihydrofluorescein (DCFH2) by cytosolic esterases, which causes the probe’s retention in the cell to allow detection of intracellular oxidizing reactions (Figure 1). In the presence of RONS, DCFH2 is oxidized to 2',7'-dichlorofluorescein (DCF) that serves as the fluorescent indicator and can be easily determined, providing a simple and sensitive analytical method for detecting intracellular oxidants (Glebska and Koppenol, 2003). However, oxidation of DCFH2 in cellular systems reflects not only the rate of production of oxidants and the rate constants for the critical one-electron oxidation step but also other cell-specific and context-dependent aspects. For example, ubiquitous cellular antioxidants such as glutathione (Figure 2) and other protein thiols or antioxidant moieties will attenuate the generation of fluorescence, dependent on the overall redox status (Albrecht et al., 2011). Moreover, despite the popularity of DCFH2 to assess oxidative stress, the identity of oxidant species responsible remains largely unclear. The correct use and interpretation of the DCFH2 assay require knowledge about contraindications or pitfalls. Notably, DCFH2 may be a valuable and sensitive indicator of cellular toxicity, but novel generations of probes must provide information about the exact cellular localization of the oxidizing RONS, reaction rates with different types of RONS, kinetics of the metabolism or decomposition of the oxidized probe, and correlation with downstream signaling (Halliwell and Whiteman, 2004; Winterbourn, 2008; Murphy et al., 2011). Carefully attention to these facts will eliminate erroneous interpretations and help to move exciting approaches of free radical research forward.
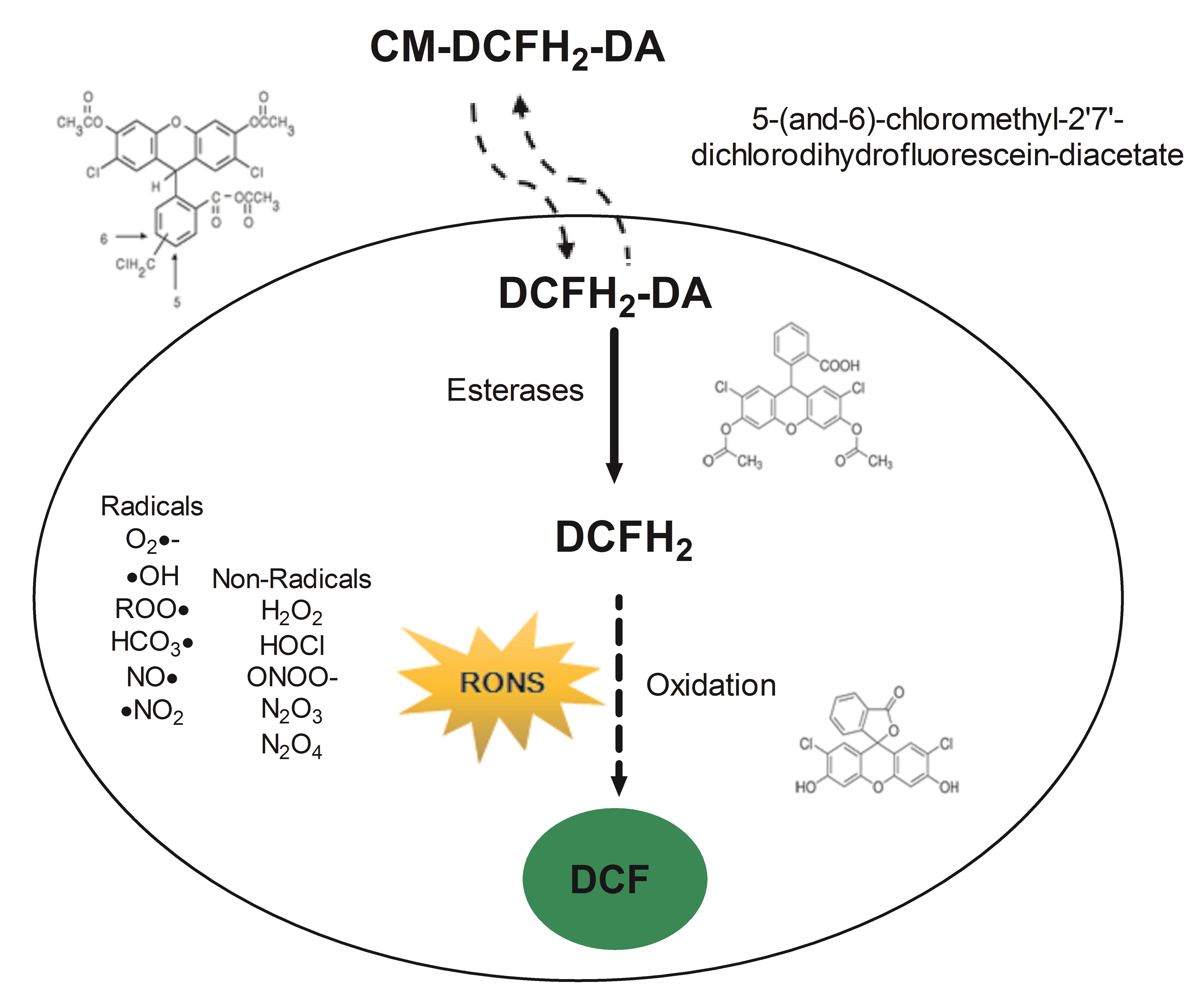
Figure 1. Schematic overview of the workflow described in this protocol. CM-DCFH2-DA diffuses into the cell, intracellular esterases hydrolyze the acetate groups, generating 2',7'- dichlorodihydrofluorescein (DCFH2) that reacts with intracellular oxidants resulting in the observable highly fluorescent compound 2',7'- dichlorofluorescein (DCF).
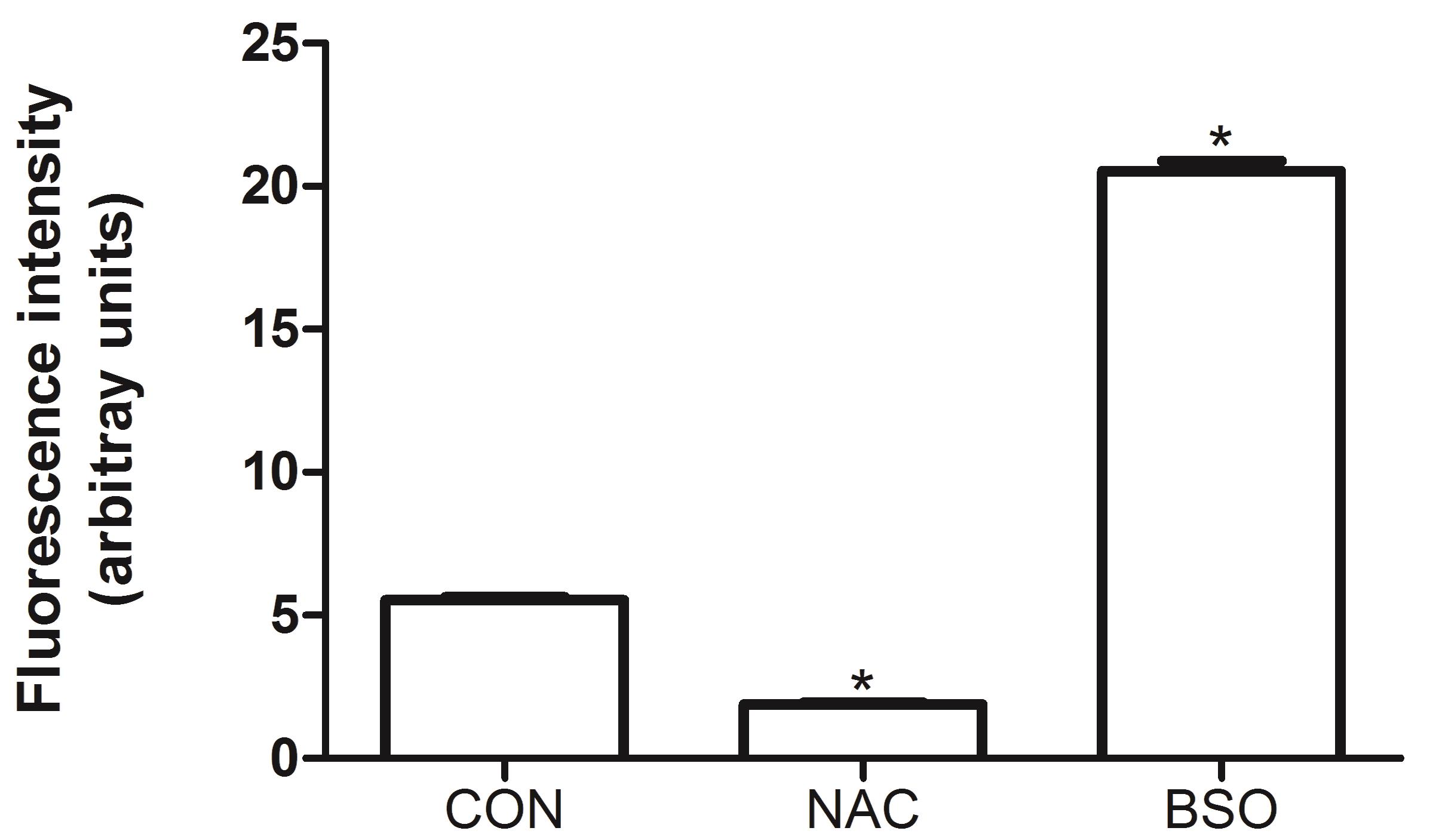
Figure 2. Effect of N-acetylcysteine (NAC) and buthionine sulfoximine (BSO) on the formation of DCF. The antioxidant NAC (5 mM) and the pro-oxidant BSO (10 µM), which significantly deplete total glutathione levels by inhibiting γ-glutamyl-cysteine synthetase (a key enzyme in glutathione biosynthesis), were evaluated on human umbilical vein endothelial cells (HUVEC) showing extremes of low and high oxidation of DCFH2, respectively. HUVEC cells were seeded at the density of 2.0 × 106 cells/cm2 for 24 h. BSO and NAC were preincubated for 1 h. The oxidation of DCFH2 in NAC or BSO treated HUVEC was compared with that of untreated control (CON) cells. The measurements of fluorescence were executed at room temperature. Data were analyzed by one-way analysis of variance with Dunnett’s Multiple Comparison Test. The data represent the means ± SEM. *P < 0.05 versus CON.
Materials and Reagents
CM-DCFH2-DA (Invitrogen Life Technologies/Molecular Probes, EUA), storage temperature at -20°C
High quality anhydrous dimethylsulfoxide (DMSO), dimethyl-formamide (DMF) or 100% ethanol, H2O2, and organic salts (Merck, Brazil), storage in 15/30°C
Loading buffer, such as a simple physiological buffer (PBS), storage in 4°C fridge
N-acetyl-L-cysteine (NAC) (Sigma, catalog number: A7250), storage in 4°C fridge
Buthionine Sulfoximine (BSO) (Sigma, catalog number: B2515), storage in 4°C fridge
HUVEC were obtained from the American Type Culture Collection, cultivated in RPMI 1640 medium supplemented with 10% fetal bovine serum, penicillin, and streptomycin antibiotics. Experiments were performed at 37°C and in a humidified atmosphere of 95% air and 5% CO2.
PBS (see Recipes)
Phosphate buffer (see Recipes), storage in 4°C fridge
Equipment
Spectrophotometer model U-2010 (Software UV Solutions, version 1.1, Hitachi, Japan)
96-well microplates (black-walled, clear bottom; Corning, catalog number: CLS3829)
Software
GraphPad Prism Version 5.01 (or higher) (GraphPad Software Inc.)
Procedure
Remove growth media from adherent or suspended cells and then wash twice with PBS 1×. Balance to 37°C before use. The 1× Buffer can be kept frozen or at 4°C for future use.
The medium should not contain phenol red or other colorimetric dyes, before and during the assay. The CM-DCFH2-DA should only be dissolved in anhydrous DMSO, DMF, or ethanol (100%). The working solutions must be freshly prepared in a light-protected vial, and tightly closed until use.
For a standard assay used often, dissolve 50 mg of CM-DCFH2-DA (MW 487.3) in 2.565 ml of DMSO or absolute ethanol (40 mM). Aliquots can be kept at -20°C for at least 3 months. Dilute 100 µl of the stock solution with 39.9 ml of pre-warmed culture medium (final working concentration 100 μM), vortex, and add to the cell culture wells.
CM-DCFH2-DA should be utilized in concentrations as low as possible to decrease potential artifacts, such as incomplete hydrolysis, and contamination by hydrolytic by-products. The ideal working solution for each application must be determined empirically, to obtain a good signal-to-noise ratio, e.g., by 2-3 further 2-fold dilutions of the standard concentration.
Incubate the cells in the Ultra-Low Cluster Plate (specific black coated 96-well dish) in the cell incubator (37°C), with high relative humidity (95%), and a controlled level of carbon dioxide (5%) in the dark. Usually, a loading time of 2 h is sufficient. Additionally, if multi-well plates and an automated plate reader are used, it is crucial to estimate the total amount of cells, cell density, and/or total protein level for each individual well, to allow normalization and secure optimal cell density and viability.
Wash carefully with the cell buffer, followed by incubation, allowing a brief recovery period for cellular esterases to hydrolyze the acetate groups. Let the dye respond to oxidation.
A suggested positive buffer control is H2O2, or tert-Butyl hydroperoxide (TBHP) at a final concentration of 50 μM.
The redox state of the sample can be measured via the increasing fluorescence of DCF in cells, as measured at 530 nm when the sample is excited at 485 nm.
Importantly, the fluorescence of negative controls without DCFH2-DA maintained in loading buffer needs to be determined as background.
In most healthy cells, oxygen radicals are eliminated by cellular enzymes and/or natural antioxidants, resulting in a usually low level of fluorescence.
Power up the fluorescence plate reader at a suitable interval prior to the initiation of the time course measurements. Describe the plate dimension situations and choose the appropriate filter set for excitation/detection of fluorescence.
Read the fluorescence on each plate and print the plate data or save in digital form.
Average the interpretations from the triplicates for each experimental condition and calculate the percent coefficient of variation (CV) for each of the means.
Subtract the fluorescence average of the cell-free treatment control from that of the condition treated under the equivalent experimental conditions to determine the intracellular DCF fluorescence. Calculate the mean fluorescence and CV for each triplicate, confirming that the triplicates are consistent.
Data analysis
Techniques for measuring RONS with the DCFH2-DA/DCFH2 system have certain weaknesses, since all the assays are prone to numerous artifacts, resulting from sample preparation or from the analytical method itself. Organic solvents, such as DMSO or ethanol, which are used to dissolve the test compounds, are another difficulty that all tests have in common. DMSO and ethanol are powerful •OH scavengers, which—although they are highly diluted in the assay and mainly extracellular—could lead to an underestimation of the amount of RONS produced. Therefore, appropriate controls are very important, because the results can be altered by various reactions taking place in the culture medium, or by handling of the cell cultures that may lead to oxidative stress (Tarpey et al., 2004). RONS measurements in biological systems do not provide absolute quantification of RONS levels, as discussed below (Figure 3).
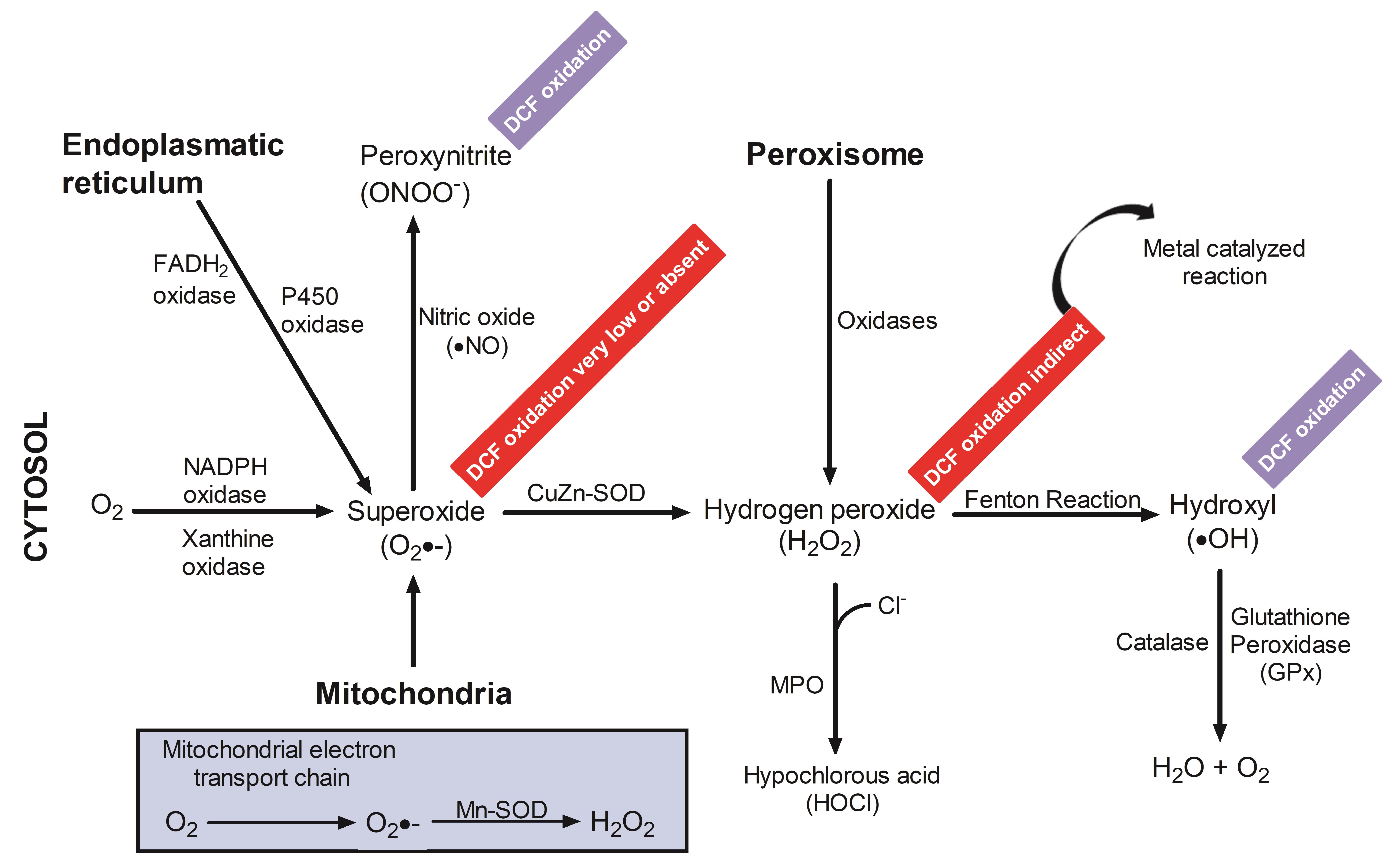
Figure 3. Schematic representation of intracellular pathways and rates of DCFH 2reactions with RONS. DCFH2 has been utilized as an indicator of the respiratory burst in macrophages and neutrophils, as well as to estimate oxidant production in response to several stimulations in other abundant cell types. However, detection of DCF to situations under which the reactive species oxidize DCFH2 may be indicative of •OH, ONOO–, but O2•−, and H2O2 alone are not.
Notes
DCFH2 has little or no reactivity with O2•–
O2•–, produced by the xanthine/XO system, is quickly inhibited by superoxide dismutase, which prevents detection by oxidation of DCFH2 (LeBel et al., 1992).
Reduction of cytochrome c induced by potassium O2•– does not rise DCF fluorescence, indicating that O2•– does not directly oxidize DCFH2 (Zhu et al., 1994).
A steady-state radiolysis providing a continuous supply of O2•– also does not cause significant oxidation of DCFH2 (Zhu et al., 1994; Wrona et al., 2005).
H2O2 cannot directly oxidize DCFH2
H2O2-dependent oxidation of DCFH2 to DCF occurs slowly, if at all, in the absence of ferrous iron. However, DCF formation is greatly enhanced in the presence of heme-containing substances, such as hematin, peroxidases, cytochrome c, or other redox-active metal ions (Cathcart et al., 1984; Kalyanaraman et al., 2012).
Peroxidases are capable of inducing DCFH2 oxidation in the absence of H2O2 (Rota et al., 1999). Therefore, increases in cellular peroxidase or heme activity are likely to be equally, if not more important than H2O2, in determining rates of DCF formation and cellular fluorescence (Ohashi et al., 2002).
Enhanced intracellular iron trafficking, such as transferrin receptor resulting in increased iron uptake into endothelial cells, usually promotes H2O2-dependent DCF fluorescence (Tampo et al., 2003).
Oxidation of DCFH2 by the •OH radical
For the Fe2+/H2O2 system (Fenton reaction), •OH scavengers such as DMSO, ethanol, mannitol, and Tris only partially inhibit DCFH2 oxidation (LeBel et al., 1992).
Free •OH, which is very short lived (<20 ms in a typical physiological environment), is not involved in the oxidation of DCFH2, while site-specific •OH formed by the binding of the DCFH2 carboxyl group to Fe2+ can oxidize DCFH2. Thus, Fe2+/H2O2–induced •OH generation (indicated by the site specific oxidation of salicylic acid to 2,3-dihydroxybenzoic acid), which is proportional to DCFH2 oxidation, can be repressed by DMSO and mannitol (Zhu et al., 1994).
DMSO has been observed to inhibit DCFH2 oxidation in several cells or cell lines, where •OH has also been shown to oxidize DCFH2 with high efficacy (Wrona et al., 2005).
DCFH2 as an excellent indicator of ONOO– formation in living cells
ONOO–-mediated oxidation of DCFH2 is much more rapid and efficient than oxidation by H2O2, NO, or O2•–. ONOO– reacting with DCFH2 reached a steady-state after 2 min, whereas H2O2 and NO continued to oxidize DCFH2 for at least 10 min (Possel et al., 1997).
In vitro oxidation of DCFH2 mediated by ONOO– was not inhibited by •OH scavengers, and was not dependent upon metal ion-catalyzed reactions. DCFH2-DA was not susceptible to oxidation by ONOO–, while ONOO– could readily oxidize DCFH2 (Crow, 1997). The lack of an inhibitory effect by the •OH scavenger mannitol and DMSO indicates that the oxidation reaction of ONOO– with DCFH2 is not based on the formation of free H2O2, which rapidly forms from the combination of two •OH radicals.
Metal catalysis is not required for efficient ONOO–-mediated DCFH2 oxidation. This reaction appears to be mediated directly by ONOO–, and does not require the secondary formation of other free radicals (Kooy et al., 1997).
HKGreen-1 possesses a ketone unit linked to a DCF moiety through an aryl ether linkage and provided a highly selective fluorescent probe for the detection of ONOO– in primary cultured neuronal cells (Yang et al., 2006).
Recipes
PBS
10× Buffer. Prepare 1× Buffer by diluting 10× buffer in deionized water to make 100 ml 1× Buffer. Combine 10 ml 10× Buffer with 90 ml deionized water and mixture gently. Marker this solution as “1× Buffer.”
Phosphate buffer (10 mM, pH 7.4) to lyse cells
5 ml of cold absolute methanol (v/v)
10 µl of Triton X100
5 ml of Buffer A + B
A: 9.5 ml 200 mM KH2PO4
B: 40.5 ml 200 mM K2HPO4
Acknowledgments
DS receives project-related support from the EU Horizon 2020 under grant agreement n. 634413 (EPoS, European Project on Steatohepatitis) and 777377 (LITMUS, Liver Investigation on Marker Utility in Steatohepatitis), and by the German Research Foundation collaborative research project grants DFG CRC 1066/B3 and CRC 1292/B8.
Competing interests
The authors declare they have no competing interests.
References
- Albrecht, S. C., Barata, A. G., Grosshans, J., Teleman, A. A. and Dick, T. P. (2011). In vivo mapping of hydrogen peroxide and oxidized glutathione reveals chemical and regional specificity of redox homeostasis. Cell Metab 14(6): 819-829.
- Brandes, R. P., Rezende, F. and Schröder, K. (2018). Redox Regulation Beyond ROS: Why ROS Should Not Be Measured as Often. Circ Res 123(3): 326-328.
- Cathcart, R., Schwiers, E. and Ames, B. N. (1984). Detection of picomole levels of lipid hydroperoxides using a dichlorofluorescein fluorescent assay. Methods Enzymol 105: 352-358.
- Crow, J. P. (1997). Dichlorodihydrofluorescein and dihydrorhodamine 123 are sensitive indicators of peroxynitrite in vitro: implications for intracellular measurement of reactive nitrogen and oxygen species. Nitric Oxide 1(2): 145-157.
- Dröge, W. (2002). Free radicals in the physiological control of cell function. Physiol Rev 82(1): 47-95.
- Dikalov, S. I. and Harrison, D. G. (2014). Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid Redox Signal 20(2): 372-382.
- Glebska, J. and Koppenol, W. H. (2003). Peroxynitrite-mediated oxidation of dichlorodihydrofluorescein and dihydrorhodamine. Free Radic Biol Med 35(6): 676-682.
- Halliwell, B. and Whiteman, M. (2004). Measuring reactive species and oxidative damage in vivo and in cell culture: how should you do it and what do the results mean? Br J Pharmacol 142(2): 231-255.
- Halliwell, B. (2012). Free radicals and antioxidants: updating a personal view. Nutr Rev 70(5): 257-265.
- Harris, I. S. and Brugge, J. S. (2015). Cancer: The enemy of my enemy is my friend. Nature 527(7577): 170-171.
- Hempel, S. L., Buettner, G. R., O'Malley, Y. Q., Wessels, D. A. and Flaherty, D. M. (1999). Dihydrofluorescein diacetate is superior for detecting intracellular oxidants: comparison with 2',7'-dichlorodihydrofluorescein diacetate, 5(and 6)-carboxy-2',7'-dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123. Free Radic Biol Med 27(1-2): 146-159.
- Kalyanaraman, B., Darley-Usmar, V., Davies, K. J., Dennery, P. A., Forman, H. J., Grisham, M. B., Mann, G. E., Moore, K., Roberts, L. J., 2nd and Ischiropoulos, H. (2012). Measuring reactive oxygen and nitrogen species with fluorescent probes: challenges and limitations. Free Radic Biol Med 52(1): 1-6.
- Kooy, N. W., Royall, J. A. and Ischiropoulos, H. (1997). Oxidation of 2',7'-dichlorofluorescin by peroxynitrite. Free Radic Res 27(3): 245-254.
- LeBel, C. P., Ischiropoulos, H. and Bondy, S. C. (1992). Evaluation of the probe 2',7'-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem Res Toxicol 5(2): 227-231.
- Murphy, M. P., Holmgren, A., Larsson, N. G., Halliwell, B., Chang, C. J., Kalyanaraman, B., Rhee, S. G., Thornalley, P. J., Partridge, L., Gems, D., et al. (2011). Unraveling the biological roles of reactive oxygen species. Cell Metab 13(4): 361-366.
- Ohashi, T., Mizutani, A., Murakami, A., Kojo, S., Ishii, T. and Taketani, S. (2002). Rapid oxidation of dichlorodihydrofluorescin with heme and hemoproteins: formation of the fluorescein is independent of the generation of reactive oxygen species. FEBS Lett 511(1-3): 21-27.
- Possel, H., Noack, H., Augustin, W., Keilhoff, G. and Wolf, G. (1997). 2,7-Dihydrodichlorofluorescein diacetate as a fluorescent marker for peroxynitrite formation. FEBS Lett 416(2): 175-178.
- Rota, C., Fann, Y. C. and Mason, R. P. (1999). Phenoxyl free radical formation during the oxidation of the fluorescent dye 2',7'-dichlorofluorescein by horseradish peroxidase. Possible consequences for oxidative stress measurements. J Biol Chem 274(40): 28161-28168.
- Tampo, Y., Kotamraju, S., Chitambar, C. R., Kalivendi, S. V., Keszler, A., Joseph, J. and Kalyanaraman, B. (2003). Oxidative stress-induced iron signaling is responsible for peroxide-dependent oxidation of dichlorodihydrofluorescein in endothelial cells: role of transferrin receptor-dependent iron uptake in apoptosis. Circ Res 92(1): 56-63.
- Tarpey, M. M., Wink, D. A. and Grisham, M. B. (2004). Methods for detection of reactive metabolites of oxygen and nitrogen: in vitro and in vivo considerations. Am J Physiol Regul Integr Comp Physiol 286(3): R431-444.
- Winterbourn, C. C. (2008). Reconciling the chemistry and biology of reactive oxygen species. Nat Chem Biol 4(5): 278-286.
- Wrona, M., Patel, K. and Wardman, P. (2005). Reactivity of 2',7'-dichlorodihydrofluorescein and dihydrorhodamine 123 and their oxidized forms toward carbonate, nitrogen dioxide, and hydroxyl radicals. Free Radic Biol Med 38(2): 262-270.
- Yang, D., Wang, H. L., Sun, Z. N., Chung, N. W. and Shen, J. G. (2006). A highly selective fluorescent probe for the detection and imaging of peroxynitrite in living cells. J Am Chem Soc 128(18): 6004-6005.
- Zhu, H., Bannenberg, G. L., Moldéus, P. and Shertzer, H. G. (1994). Oxidation pathways for the intracellular probe 2',7'-dichlorofluorescein. Arch Toxicol 68(9): 582-587.
Article Information
Copyright
© 2021 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Dornas, W. and Schuppan, D. (2021). Measurement of Reactive Oxygen and Nitrogen Species in Living Cells Using the Probe 2',7'-Dichlorodihydrofluorescein. Bio-protocol 11(24): e4279. DOI: 10.21769/BioProtoc.4279.
Category
Developmental Biology > Cell signaling > Stress response
Medicine
Cell Biology > Cell metabolism > Other compound
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.