- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Dictyostelium Cultivation, Transfection, Microscopy and Fractionation



Published: Vol 5, Iss 11, Jun 5, 2015 DOI: 10.21769/BioProtoc.1485 Views: 14725
Reviewed by: Arsalan DaudiAlexandros AlexandratosAnonymous reviewer(s)
Original research article
The authors used this protocol in:

May 2014
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics







Abstract
The real time visualisation of fluorescently tagged proteins in live cells using ever more sophisticated microscopes has greatly increased our understanding of the dynamics of key proteins during fundamental physiological processes such as cell locomotion, chemotaxis, cell division and membrane trafficking. In addition the fractionation of cells and isolation of organelles or known compartments can often verify any subcellular localisation and the use of tagged proteins as bait for the immunoprecipitation of material from cell fractions can identify specific binding partners and multiprotein complexes thereby helping assign a function to the tagged protein. We have successfully applied these techniques to the Dictyostelium discoideum protein TSPOON that is part of an ancient heterohexamer membrane trafficking complex (Hirst et al., 2013). TSPOON is the product of the tstD gene in Dictyostelium and is not required for growth or the developmental cycle in this organism. Dictyostelium amoebae will exist in a vegetative phase where growth is sustained by the phagocytosis of bacteria. When this food source is spent they enter a developmental phase where the amoebae aggregate, via chemotaxis to extracellular waves of cAMP, into multicellular structures that subsequently form a fruiting body containing viable spores (Muller-Taubenberger et al., 2013). In the laboratory this cycle takes less than 24 h to complete and as a further aid to manipulation the requirement for a bacterial food source has been circumvented by the derivatisation of the wild type and isolation of axenic strains that can also grow in a nutrient rich broth. Axenic strains like Ax2 are the mainstay of laboratory research using Dictyostelium (Muller-Taubenberger et al., 2013). A description of Dictyostelium cell cultivation, the generation of cell lines that overexpress TSPOON-GFP and TSPOON null cells, and subsequent analysis (Muller-Taubenberger and Ishikawa-Ankerhold, 2013) is detailed below.
Keywords: DictyosteliumMaterials and Reagents
- Dictyostelium Ax2 cells can be obtained from the Dictyostelium stock centre (http://dictybase.org/StockCenter/StockCenter.html)
Note: Ax2 strain DBS0235521 is recommended due to the relatively few duplications within its genome (Bloomfield et al., 2008). - HL5 with glucose (Formediun, catalog number: HLG0102 )
- LOFLO (Formediun, catalog number: LF1001 )
- Ultra-pure dH2O (e.g. Milli-Q system, EMD Millipore)
- KH2PO4 (Thermo Fisher Scientific, catalog number: P/4800/53 )
- K2HPO4 (anhydrous) (Sigma-Aldrich, catalog number: 60353 )
- MgSO4.7H2O (VWR International, catalog number: 25165.260 )
- CaCl2.2H2O (VWR International, catalog number: 22317.260 )
- HEPES (Sigma-Aldrich, catalog number: H4034 )
- KCl (Thermo Fisher Scientific, catalog number: P/4280/53 )
- NaCl (VWR International, catalog number: 27810.295 )
- NaHCO3 (VWR International, catalog number: 27778.236 )
- NaH2PO4.2dH2O (Thermo Fisher Scientific, catalog number: 10723621 )
- Na2HPO4 (anhydrous) (Thermo Fisher Scientific, catalog number: S4520/53 )
- KOH (VWR International, catalog number: 26668.263 )
- Trizma® base (Sigma-Aldrich, catalog number: T1503 )
- EDTA disodium salt dihydrate (VWR International, catalog number: 20302.260 )
- HCl (Thermo Fisher Scientific, catalog number: H/1200/PB08 )
- Klebsiella aerogenes (available from http://dictybase.org/StockCenter/StockCenter.html)
- 50 mM L-ascorbic acid (Sigma-Aldrich, catalog number: A5960 )
- Ethanol (Sigma-Aldrich, catalog number: 3112 )
- Chloroform (VWR International, catalog number: 22711.290 )
- Plasmids pDT51, pDT61, pDT70 and pJH101 together with gene knockout vectors for iplA, mscS, mclN, phdA and dagA can be obtained from David Traynor
Note: Plasmid pLPBLP can be ordered through dictyBase (http://www.dictybase.org/db/cgi-bin/dictyBase/SC/plasmid_details.pl?id=9). - HiSpeed® Plasmid Midi Kit (QIAGEN, catalog number: 12643 )
- Hygromycin 100 mg/ml sterile solution (Invivogen HygroGold, catalog number: ant-hg-5 )
- Blasticidin S 100 mg/ml sterile solution (Invivogen, catalog number: ant-bl-1 )
- G418 (20 mg/ml solution) (Life Technologies Geneticin®, catalog number: 11811-031 )
- Quick-gDNA MiniPrep kit (Zymo Reasearch, catalog number: D3025 )
- Expand 20 kbPLUS PCR system (dNTPack) (F. Hoffmann-La Roche, catalog number: 04743814001 )
- Oligonucleotide primers (custom synthesised by Sigma-Aldrich)
- Freezing medium [sterilised horse serum containing 7.5% (v/v) DMSO]
- Horse serum (Sigma-Aldrich, catalog number: H1270 )
- DMSO (Sigma-Aldrich, catalog number: D2650 )
- Master stock of 200 mM cAMP
- 4x NuPAGE® LDS sample buffer (Life Technologies, Novex, catalog number: NP0008 )
- Protein A (PA) sepharose CL-4B beads (GE Healthcare, catalog number: 17-0780-01 )
- Acetone (Thermo Fisher Scientific, catalog number: A/0560/17 )
- Protease inhibitor cocktail (F. Hoffmann-La Roche, catalog number: 11873580001 )
- Precast NuPAGE 4-12% Bis-Tris gels (Life Technologies, Novex)
- Coomassie G-250 SimplyBlue safe stain (Life Technologies, InvitrogenTM, catalog number: LC6060 )
- PierceTM BCA protein assay (Thermo Fisher Scientific, catalog number: 23225 )
- Phenol solution (Equilibrated with 10 mM Tris HCl, pH 8.0, 1 mM EDTA) (Sigma-Aldrich, catalog number: P4557 )
- Adenosine 3′, 5′-cyclic monophosphate (cAMP) (free acid) (Sigma-Aldrich, catalog number: A9501 )
- TritonTM X-100 solution (~10% in H2O) (Sigma-Aldrich, catalog number: 93443 )
- Sodium dodecyl sulphate (SDS) solution (10% in H2O SDS) (Sigma-Aldrich, catalog number: 71736 )
- SM agar (Formediun, catalog number: SMA0120 )
- SM broth (Formediun, catalog number: SMB0102 )
- Trichloroacetic acid (VWR International, catalog number: 102863H )
- Q5 (NEB, catalog number: M0491 )
- Pwo (Roche Diagnostics, catalog number: 11644947001 )
- KOD polymerases (Merk Millipore, catalog number: 71086 )
- Antifade mounting medium Fluoromount-G® (Cambridge Bioscience, catalog number: 0100-01 ) 71086 or Prolong® (Life Technologies, catalog number: P10144 )
- Axenic medium (see Recipes)
- LOFLO medium (see Recipes)
- SM agar (see Recipes)
- SM broth (see Recipes)
- Trichloroacetic acid (see Recipes)
- KK2 buffer (see Recipes)
- KK2C buffer (see Recipes)
- 1 M MgSO4 (see Recipes)
- 1 M CaCl2 (see Recipes)
- Electroporation buffer E50 (see Recipes)
- Dense suspension of Klebsiella aerogenes (see Recipes)
- Tris-HCl buffer (see Recipes)
- TE buffer (see Recipes)
- 20 mg/ml G418 (see Recipes)
- Freezing medium (see Recipes)
- 50 mM L-Ascorbic (see Recipes)
- Master stock of cAMP (see Recipes)
- Phosphate buffered saline (PBS) (see Recipes)
- PBS-T (see Recipes)
- Elution buffer (see Recipes)
Equipment
- Sterile tissue culture dishes [(100 x 20 mm style, Corning, catalog number: 353003 or Nunc, catalog number: 172958 ) (35 x 10 mm style, Corning, catalog number: 353001 )]
- Sterile 96 flat bottomed well tissue culture dishes (Corning Costar, catalog number: 3799 )
- Sterile 50 ml plastic tubes (SARSTEDT AG, catalog number: 62.547.004 or Corning, catalog number: 352070 )
- 1.8 ml CryoTube vials (Thermo Fisher Scientific, catalog number: 377267 )
- Pipetting reservoir (120 ml capacity) (Thermo Fisher Scientific, catalog number: 10717964 )
- Steribag pouches (140 mm x 50 mm x 330 mm) (Thermo Fisher Scientific, catalog number: 12728995 )
- Microfuge tubes (Sarstedt Micro tube 1.5 ml) (SARSTEDT AG, catalog number: 72.690 )
- Lab-Tek chambered coverglass with cover (Thermo Fisher Scientific, catalog number: 155411 and 155380 )
- Glass bottom microwell 35 mm petri dishes with 20 mm microwell (MatTek, catalog number: P35G-1.5-20-C )
- Disposable cell spreaders (Biologix, catalog number: 65-100 )
- Cryo preservation module (Agilent StratCooler Lite Preservation module, catalog number: 400006 )
- 21G (0.8 mm x 40 mm) needles (Becton Dickinson, catalog number: 304432 )
- Spatula (150 x 4 mm) (Thermo Fisher Scientific, catalog number: 11503482 )
- Platinum wire inoculating loop and insulated holder (Thermo Fisher Scientific, catalog number: 10669652 and 12882775 )
- Nunc sterile plastic needles and loops (Thermo Fisher Scientific, catalog number: 254399 and 254410 )
- Syringe filters (0.22 µm) for sterilisation of small volumes (Elkay, catalog number: E25-PS22-50S )
- Sterile disposable syringes for sterilisation of small volumes (Becton Dickinson, Plastipak 50 ml, catalog number: 30086 ; 20 ml, catalog number: 300613 ; 10 ml, catalog number: 302188 )
- Swin-LokTM plastic filter holder 25 mm (GE Healthcare, catalog number: 420200 )
- Prefilters. 25 mm cellulose absorbent pads (Merck Millipore, catalog number: AP1002500 )
- Nucleopore Track-Etch polycarbonate membrane 3.0 µm (GE Healthcare, catalog number: 110612 )
- Micro bio-spin chromatography column (Bio-Rad Laboratories, catalog number: 732-6204 )
- Duran bottles (or equivalent autoclavable media bottles) 1,000 ml, 500 ml and 150 ml (Thermo Fisher Scientific, catalog numbers: FB-800-1000 , FB-800-500 and 11699888 )
- Foam stoppers (50 mm x 50 mm and 50 mm x 38 mm) (Thermo Fisher Scientific, catalog number: 11522563 and 11512563 )
- Aluminium foil (300 mm x 75 mm x 0.02 mm) (Prowrap, catalog number: JR307518U )
- Disposable plastic filter bottle units (150 ml) (Millipore Stericup, SCGPU01RE ) or 500 ml (Millipore Stericup, catalog number: SCGPU05RE )
- Steritop bottle top filter unit (1,000 ml) (EMD Millipore, SCGPT10RE )
- Corning Filter System (1,000 ml) (Corning, catalog number: 430186 )
- Universal bottle (Thermo Fisher Scientific, catalog number: BTS-402-050M )
- Temperature controlled shaking incubator (e.g. Infors Multitron standard or Innova 4330)
- Temperature controlled incubator (e.g. Panasonic, model: MIR-254 PE or MIR-154 PE )
- Tissue culture hood (e.g. Scanlaf Mars Safety 2)
- Ultrapure water supply (e.g. Milli-Q® Advantage A10 Ultrapure Water Purification System)
- Autoclave
- Water bath (e.g. Grant SUB Aqua 12 Plus)\
- pH meter and calibration buffers (e.g. Hanna Instruments Edge Hybrid pH, catalog number: HI-2020-02 )
- Cell counter (e.g. Beckman Coulter Z1) or haemocytometer (Weber Scientific)
- Bench top centrifuge (e.g. Thermo Fisher Scientific, model: IEC CL30 or Eppendorf, model: 5702R )
- Bench top microfuge (e.g. Eppendorf, model: 5424 or 5424R )
- Thermocycler (96 x 0.2 ml tube block) (e.g. Biometra TGradeint or Applied Biosystems Veriti)
- Programmable peristaltic pump (e.g. Watson Marlow, model: 505Di )
- 8 channel electronic pipette (e.g. eLine 50-1,200 µl) (Sartorius, catalog number: 730391 ) and 50-1,200 µl tips (SafetySpaceTM filter tips) (Sartorius, catalog number: 79121F )
- Confocal microscope (e.g. ZEISS, model: LSM710 or LSM780 )
- TIRF microscope (e.g. Nikon Corporation)
Note: Both the Zeiss confocal and Nikon TIRF microscopes are supplied with the manufacturers image analysis software (Zen and NIS-Elements respectively) but due to the limited time users have on this equipment image analysis is usually performed on a dedicated computer work station running these software packages that are an additional purchase from the manufacturers. Free to use image analysis software (open source) packages are also routinely used. ImageJ and Fiji can be used for the confocal and TIRF images. ImageJ can be downloaded from http://imagej.nih.gov/ij and Fiji from http://fiji.sc/Fiji. - Electroporator (e.g. BIO-RAD Xcell) and 1 mm gap width cuvettes (Cell Projects, catalog number: EP-201 )
- Potter-Elvehjem homogensier (Wheaton, catalog number: 358049 )
- Ultracentrifuge (Beckman Coulter, model: Optima MAX-XP )
- Orbitrap Mass Spectrometer (Thermo Fisher Scientific)
- 0.2 ml PCR strip tube with 12 wells and separate cap (VWR International, catalog number: 53509-306 )
- TLA-110 rotor (Beckman Coulter)
- XCell SureLock® Mini-Cell and XCell II™ blot module (Life Technologies, Novex, catalog number: EI0002 ) and power supply (Bio-Rad Laboratories, PowerPacTMHC High-Current Power Supply, catalog number: 164-5052 )
Procedure
- Growing Dictyostelium cells
- If possible all work with live Dictyostelium discoideum cells should be done in a 22 °C temperature controlled room. The initiation and passaging of cultures either in shaken suspension or in dishes should be done under sterile conditions in a tissue culture hood. It is very important to renew stocks every month from spores or frozen stocks stored at -80 °C or in liquid nitrogen.
- Ax2 or the TSPOON null cells (HM1725 and HM1727) cells are grown in shaken suspension (22 °C, 180 rpm) in axenic medium in 250 ml or 500 ml conical flasks (seed at 0.5-2 x 105 cells/ml) and harvested in mid log phase (2-5 x 106 cells/ml). Doubling time in these conditions is normally 8-11 h.
- Alternatively, cells can be grown on tissue culture plates and harvested when confluent (each plate should yield 1-2 x 107 cells) (Figure 1). This is useful when relatively few are required for confocal or TIRF microscopy.

Figure 1. A phase contrast light photomicrograph of a near confluent lawn of Dictyostelium amoebae. The cells were grown and photographed in a 10 cm tissue culture dish containing 12 ml of axenic medium. Note that there are few gaps between individual cells. The scale is shown in the bottom right.
- If possible all work with live Dictyostelium discoideum cells should be done in a 22 °C temperature controlled room. The initiation and passaging of cultures either in shaken suspension or in dishes should be done under sterile conditions in a tissue culture hood. It is very important to renew stocks every month from spores or frozen stocks stored at -80 °C or in liquid nitrogen.
- Electroporation of Dictyostelium cells (based on Pang et al., 1999)
- Harvest mid log phase cells into 50 ml sterile plastic tubes and centrifuge at 300 x g for 2-3 min to pellet the cells. Aspirate supernatant and add 50 ml of ice cold E50 buffer and resuspend the cell pellet by gently tapping and shaking the tube.
- Determine the number of cells/ml using an automated cell counter or a haemocytometer.
- Centrifuge at 300 x g for 2-3 min to pellet the cells and then resuspend at 4 x 107/ml in ice cold E50 buffer. Place the cell suspension on ice and incubate for 5 min (if the cells start to settle then gently mix by agitation).
- For overexpression plasmids such as pJH101 (TSPOON-GFP fusion driven by the constitutively active actin15 promoter) or pDT58 and pDT61 (both contain the TSPOON-GFP fusion driven by the TSPOON promoter), transfer 0.1 ml of the cell suspension to a pre-chilled sterile electroporation cuvette (1 mm gap width) and add 10-30 µg of supercoiled plasmid DNA (preferably in ≤15 µl of Tris-HCl or TE buffer and prepared according to the manufacturer’s instructions using the HiSpeed® Plasmid Midi Kit) then mix by gently pipetting up and down avoiding introducing air bubbles to the cell suspension. Return the cuvette to ice (Figure 2).
- The most common and reliable gene disruption in Dictyostelium is achieved with blasticidin S as the selective agent using a plasmid containing the resistance cassette (such as pLPBLP) flanked on either side with DNA homologous to the targeted gene (Faix et al., 2004; Faix and Kimmel, 2006). This targeting/disruption cassette should be freed from the remainder of the construct by complete digestion with restriction enzymes. To knockout TSPOON, plasmid pDT70 (based on pLPBLP) was cut with ApaI and SacII (these enzymes generate 3’ overhangs in the cut DNA but enzymes that leave 5’ overahangs e.g. BamHI or blunt cutters like PvuII can be used in any combination to liberate the disruption cassette). The enzymes where removed by a single phenol extraction followed by three extractions with chloroform and the cut DNA precipitated with ethanol (Sambrook and Russel, 2001). Resuspend the cut DNA at 2-3 mg/ml in Tris-HCl or TE buffer. Add 15-20 µg of the disruption cassette DNA to each 0.1 ml of cell suspension, again in a chilled sterile electroporation cuvette (1 mm gap width) and mix as in step B4a.

Figure 2. A photograph of the electroporation apparatus described in this protocol. In the background the BioRad Xcell is shown with the pod open and loaded with an electroporation cuvette (1cm gap width). In the ice bucket is two more cuvettes awaiting electroporation. In the foreground a tube (15 ml) containing the axenic medium that will be added (0.5 ml) after each cuvette is pulsed twice to electroporate the cell suspension within. A tissue culture dish with 6 wells each of which contains 3 ml of axenic medium is also shown and after recovery on ice for 5 min, 100 µls of the electroporated cell suspension will be added to each well of this dish.
- For overexpression plasmids such as pJH101 (TSPOON-GFP fusion driven by the constitutively active actin15 promoter) or pDT58 and pDT61 (both contain the TSPOON-GFP fusion driven by the TSPOON promoter), transfer 0.1 ml of the cell suspension to a pre-chilled sterile electroporation cuvette (1 mm gap width) and add 10-30 µg of supercoiled plasmid DNA (preferably in ≤15 µl of Tris-HCl or TE buffer and prepared according to the manufacturer’s instructions using the HiSpeed® Plasmid Midi Kit) then mix by gently pipetting up and down avoiding introducing air bubbles to the cell suspension. Return the cuvette to ice (Figure 2).
- The cell/DNA suspension should be electroporated quickly to minimise cell sedimentation in the cuvette.
- Remove the cuvette from the ice and dry off any excess moisture on the electrodes with a tissue before placing in the electroporation pod.
- The parameters should be set at 0.75-0.85 kV, 25 µF capacitance with resistance set at infinity (∞) for the Bio-Rad Xcell. The same parameters are used on the Bio-Rad GenePulser and GenePulserII however they may need to be optimised with electroporation devices from other manufacturers.
- Deliver two pulses 5 sec apart to each cuvette and then quickly add 0.5 ml of sterile room temperature axenic medium, mix by gentle pipetting up and down and then return the cuvette to ice for 5 min.
- For the selection process and screening of transformants see Faix et al. (2004); Kimmel and Faix (2006); Muller-Taubeberger et al. (2006).
- Remove the cuvette from the ice and dry off any excess moisture on the electrodes with a tissue before placing in the electroporation pod.
- Harvest mid log phase cells into 50 ml sterile plastic tubes and centrifuge at 300 x g for 2-3 min to pellet the cells. Aspirate supernatant and add 50 ml of ice cold E50 buffer and resuspend the cell pellet by gently tapping and shaking the tube.
- Isolation of overexpression and knockout cell lines
- For each cuvette prepare a 6 well tissue culture plate with 3 ml of axenic medium in each well.
- Remove a cuvette from ice and resuspend the cells by gentle pipetting before adding 0.1 ml to each well of the tissue culture dish.
- Incubate at 22 °C overnight (normally 16-24 h) before adding, in duplicate wells, 10 µg/ml, 20 µg/ml and 40 µg/ml G418 (for plasmids pJH101 and pDT61).
- For selection with hygromycin, add 30 µg/ml, 60 µg/ml and 90 µg/ml (for pDT58).
- Change the selection medium by aspiration to remove dead cells every 2-3 days and the wells should be confluent after 9-14 days. (Hygromycin appears to be less efficient at killing Dictyostelium amoebae than G418 and it is therefore important to replace the medium every 2-3 days as directed to ensure the removal of dead and dying cells that will become detached from the tissue culture plate).
- The wells are screened for optimum expression by dislodging the cells by pipetting up and down and removing 0.3 ml into a single well of a chambered coverslip (8 well).
- The cells are allowed to settle and attach for 20-30 min before the medium in each well is aspirated and replaced by 0.3 ml of LOFLO medium or KK2C.
- The cells are then examined by confocal or TIRF microscopy for the presence and distribution of the GFP fusion protein (Figures 3 and 4).

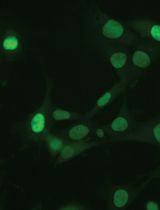
Figure 3. Overexpression of TSPOON-GFP. The top panel are cells transfected with pDT61 (TSPOON-GFP fusion protein driven by the Tspoon promoter) and selected for 14 days in axenic medium containing 10µg/ml G418. Most cells express the fusion protein but at varying levels. The bottom panel is mock transfected cells. The cells in both panels where transferred to LoFlo medium for 30 minutes before being imaged by confocal microscopy using exactly the same image collection parameters. Note the faint autofluorescence in the mock transfected cells visible as puncta. The scale is indicated in the bottom right of the figure.
Figure 4. A TIRF image of TSPOON-GFP. The amoebae were transfected and selected as in Figure 3. Cells were transferred into KK2C (plus 50 µg/ml L-ascorbic acid) 3 h prior to image collection. The scale is indicated. - To make a permanent stock, the well(s) with the optimum expression are harvested by pipetting the axenic medium up and down to dislodge the cells which are then used to initiate a larger culture (grown with the appropriate levels of G418 or hygromycin) either in shaken suspension or several 10 cm tissue culture dishes each with 12 ml of axenic medium.
- A minimum of 8 x 107 cells should be harvested and pelleted by centrifugation (300 x g for 2 min) then resuspended in 1.6 ml of freezing medium and processed as in step C23.
- To isolate knockout mutants the cell suspension should be diluted to appropriate densities to ensure clonality over a range of cell survival. This is particularly important when the targeted gene affects growth because wild type non-homologous recombinants are likely then to outgrow such a knockout, making the screen and its isolation more difficult.
- To ensure clonality, the cell suspension from a single cuvette is first added to a total of 15 ml of axenic medium in 50 ml plastic tube so that the cells are approximately 2.67 x 105/ml.
- This cell suspension is further diluted 10 (2.67 x 104/ml), 50 (5.33 x 103/ml), 100 (2.67 x 103/ml), 200 (1.33 x 103/ml) and 400 fold (0.67 x 103/ml). Each dilution is made into a total of 15 ml of axenic medium and 100 µl/well is dispensed into a separate 96 well tissue culture plate for each dilution.
- Normally 4 x 96 well plates are prepared for the 100, 200 and 400 fold dilutions so 50 ml of diluted cells should be prepared for each of these dilutions.
- Incubate the plates in a moist atmosphere (such as in a large plastic cake storage box lined with wet tissues) at 22 °C overnight (normally 16-24 h) before adding 100 µl of axenic medium containing 20 µg/ml blasticidin S to each well (10 µg/ml final).
- Change the selection medium to remove dead cells every 2-3 days by forcefully ‘throwing’ the medium into a large tissue lined (stops splash back into wells) container. Add 200 µl of fresh axenic medium containing 10 µg/ml of blasticidin S to each well.
Note: Plating out the cells into the 96 well plates and replenishing the medium is best achieved with a 8 channel electronic pipette fitted with tips each with a capacity of 1,200 µl. The cell suspension or fresh media is dispensed from a sterile plastic reservoir. These reservoirs can be reused after autoclaving in a steribag pouch. - The 96 well plate containing the least diluted cells should have confluent wells after 9 days whereas the plates containing the 100, 200 and 400 fold dilutions may take up to 21 days (times are average based on the use of many different knockout vectors but deletion of a gene that impairs growth in axenic medium is likely to increase these times). Dilutions are usually screened from plates where <30 wells/plate are occupied, as these are likely to be clonal.
- The cells from each confluent well are harvested by pipetting up and down, then transferred to 1.5 ml microcentrifuge tube containing 0.75 ml of sterile KK2 (this is to dilute residual axenic medium after the cells are pelleted).
- Refill the well with 200 µl of fresh axenic medium containing 10 µg/ml of blasticidin S. The cells are pelleted in a benchtop microfuge at full speed for 20 sec and the supernatant removed by aspiration.
- Genomic DNA from the cell pellet can then be isolated using a variety of commercial kits but we prefer the Quick-gDNA MiniPrep kit from Zymo Reasearch as it is quick, has few steps and reproducibly yields good quality genomic DNA for screening by PCR using a variety of polymerases [Expand 20 kb plus polymerase (Roche) was found to be optimal for the TSPOON screen but we have also used Q5, Pwo and KOD polymerases].
- In the case of the TSPOON knockout, isolation and screening took 17 days from the start of selection until confluent wells where observed in the plates containing the 200 and 400 fold dilutions.
- The PCR screen used TSPOON specific primer (TCP) oligonucleotides TCP15: (5’-GATGAAATTTATCAGATATTGATTTCATGAATGTTTCACC-3’) and TCP16 (5’-CTGCTGATGTTGGTTTATAGGTGGCAAACCACCATC-3’) both of which bind outside the disruption cassette to minimise false positives generated by single crossover recombination events. The gene encoding TSPOON (tstD, DDB_G0350235) is located on chromosome 1 and is flanked by genes DDB_G0271002 and DDB_G026990. The exact genomic location of TCP15 is 1:3986383..3986422 and TCP16 is 1:3982321..3982356. All the oligonucleotide primers used in this protocol are located within these coordinates.
- The PCR reactions (in 0.2 ml tubes) consisted of a total of 25 µl containing 1x Expand 20kbPLUS buffer, 0.4 nM of each primer, 5 µl of the Quick-gDNA miniprep, 500 µM dNTPs and 0.25 µl (5 units) of Expand 20kbPLUS polymerase.
- The cycling parameters were step (1) 92 °C for 1 min 15 sec; (2) 92 °C for 30 sec; (3) 52 °C for 45 sec; (4) 62 °C for 7 min; (5) 62 °C for 5 min. Steps 2 to 4 were repeated a further 36 times before the reactions were held at 4 °C until analysed by gel electrophoresis (Sambrook and Russel, 2001).
- The PCR product from wild-type clones is ~4.1 kb and TSPOON knockout clones ~4.7 kb.
- The insertion of blasticidin resistance cassette and flanking DNA introduces restriction sites into the knockout PCR product that are absent in the wild type. This is a useful further proof of correct targeting as digestion of these products with a restriction enzyme such as SmaI for TSPOON, leaves the wild type product intact but cuts the knockout product into 3 fragments.
- A total of 46 wells where screened by this method, 37 were TSPOON knockouts, 6 were wild type and 3 were not clonal as they contained both PCR products. Therefore, the targeting frequency for this the tstD gene was 80% and though high was in keeping with other genes targeted by this procedure in our laboratory such as 76% for iplA, 67% mscS, 57% mclN, 50% phdA and 12% dagA.
- The PCR reactions (in 0.2 ml tubes) consisted of a total of 25 µl containing 1x Expand 20kbPLUS buffer, 0.4 nM of each primer, 5 µl of the Quick-gDNA miniprep, 500 µM dNTPs and 0.25 µl (5 units) of Expand 20kbPLUS polymerase.
- Four TSPOON knockouts, isolated from different 96 well plates, were processed further (2 for study and 2 as backup).
- The wells containing these cells were harvested as before, the cells diluted in KK2 to 0.6-1.2 cells/µl (a confluent well contains ~105 cells).
- SM agar plates were prepared each with a 0.4 ml drop of a dense suspension of Klebsiella aerogenes bacteria (Figure 5).

Figure 5. A SM agar plate covered in a lawn of Klebsiella aerogenes with Dictyostelium colonies visible. The translucent periphery of a colony consists of vegetative amoebae while at the centre fruiting bodies and other multicellular structures have formed. Colonies suitable for harvesting and freezing should be ~1 to 1.5 cm in diameter and clearly separated from neighbours. Suitable colonies in this example are indicated by the black arrowheads. Approximately 6 amoebae in 10 µl of KK2 were directly added a 400 µl drop of a dense suspension of Klebsiella aerogenes already on the plate and then evenly spread. Colonies were visible 4 days after plating and this photograph was taken after 6 days. The scale is shown in the bottom right. - Add 10 µl of the diluted cell suspension to this drop and spread evenly over the plate with a sterile plastic spreader.
- Discrete colonies appear within 4-5 days and one per knockout was harvested when they had reached ~1.5 cm in diameter.
- The flat end of a sterilised small metal spatula was used to transfer the entire colony directly into 1.6 ml of freezing medium within a CryoTube on ice. The CryoTubes were intermittently vortexed to ensure an even suspension and then transferred to cryo preservation module chilled to 4 °C.
- The module is then transferred to a -80 °C freezer overnight and then the tubes can remain at -80 °C (cells remain viable for >2 years) or be transferred to a liquid nitrogen storage tank (indefinite viability).
- The wells containing these cells were harvested as before, the cells diluted in KK2 to 0.6-1.2 cells/µl (a confluent well contains ~105 cells).
- For each cuvette prepare a 6 well tissue culture plate with 3 ml of axenic medium in each well.
- Microscopy
- Fixed and live cells were analysed my microscopy at the vegetative or the aggregation competent stage of development.
For live vegetative cells- Simply harvest growing cells from axenic culture and transfer an appropriate number of cells so that final density is 0.5-1.5 x 105/cm2 into a glass bottom dish or chambered coverslip containing LOFLO medium (this has been formulated to minimise background and cellular autofluorescence compared to standard axenic medium and it will keep the cells vegetative for a while) or KK2C (they will start to develop in this buffer but autofluorescence and photosensitivity will be minimised).
- Incubate at 22 °C for 10 min to allow the cells to attach then aspirate the medium (do not allow the cells to dry) and replace with fresh LOFLO or KK2C. They can then be used for live cell imaging or fixed.
For developed cells- Transfer 0.5-1.5 x 105/cm2 into KK2C and replace the KK2C once the cells have attached.
- Incubate at 22 °C for 8-10 h to allow the cells to become aggregation competent.
- Alternatively transfer 1 x 106 cells into a 35 mm tissue culture dish containing 2 ml of KK2C, allow the cells to attach and then replace the KK2C and incubate at 22 °C for 1 h.
- Transfer the dish to 15 °C overnight (15-17 h) and then return the dish to 22 °C for 1 h before transfer (at 0.5-1.5 x 105/cm2) to a glass bottom dish (the cells should aggregate within 2-3 h at 22 °C).
- If larger quantities of developed cells are needed then resuspend vegetative cells at 2 x 107/ml in KK2C and transfer them into a conical flask with a volume of at least 5 times that of the cell suspension to ensure optimum aeration. The cells are shaken at 180 rpm at 22 °C for 1 h then a 100 µl droplet of KK2C, containing enough cAMP (diluted from a master stock) so that the final concentration of the entire suspension is 100 nM, is dispensed into the suspension every six minutes for 3.5-5.0 h via a programmable peristaltic pump. This procedure mimics the pulsatile waves of cAMP that are emitted through a developing population of amoebae and is useful for mutant strains defective in early development and cAMP signalling.
- Harvest the cells and pellet by centrifugation 300 x g for 2 min, aspirate off the supernatant and resuspend the cells at 1 x 107/ml in KK2C and plate as before at 0.5-1.5 x 105/cm2.
- There are numerous ways to fix Dictyostelium amoebae for immunocytochemistry (ICC) and they can be found elsewhere (DictyBase1 and 2, Jungbluth et al., 1994; Hagedorn et al., 2006).
- Simply harvest growing cells from axenic culture and transfer an appropriate number of cells so that final density is 0.5-1.5 x 105/cm2 into a glass bottom dish or chambered coverslip containing LOFLO medium (this has been formulated to minimise background and cellular autofluorescence compared to standard axenic medium and it will keep the cells vegetative for a while) or KK2C (they will start to develop in this buffer but autofluorescence and photosensitivity will be minimised).
- Confocal laser scanning microscopy.
- Live cell imaging is normally a balance between detecting the tagged fusion protein (signal to noise ratio) and damaging the cells with the laser due to free radicals produced by the illumination interacting with cellular constituents or the fluorophore tag. With a 10x or 20x lens this is never normally a problem but at these magnifications there is little or no spatial resolution within a cell to be gained given that Dictyostelium cells are ~10-15 µm in diameter.
- Normally a 63x or 100x lens is needed to resolve the subcellular localisation of a tagged protein within a Dictyostelium amoeba.
- Vegetative amoebae are particularly sensitive to damage especially from shorter wavelengths such as the 405 nm laser. An Argon laser (25-35 mW) is fitted to most confocal miscroscopes for imaging YFP, GFP and CFP fusion proteins and excessive power will result in phototoxicity and photobleaching so the laser strength should be kept to a minimum (normally 2-10% for the 488 nm laser line).
- Longer wavelength laser lines such as 561 mn used for RFP tagged proteins present less of a problem and can be used at higher power (>15%) signal can be increased by opening the pinhole.
- Avoid line averaging, slow scans and real time deconvolution.
- Developed cells are usually less sensitive to laser damage but it can be fusion protein dependent. For instance a GFP tagged protein that resides in the plasma membrane may render the cells more sensitive (due to lipid oxidation in the membrane) than a cytoplasmic protein. L-Ascorbic acid (50-100 µm final) to scavenge free radicals can be added to minimise this problem and the vitamin E analogue Trolox C has also been used to this end (add 1 mM to the growth medium and incubate overnight before imaging).
- If phototoxicity cannot be overcome then consider switching to a spinning disc confocal microscope (SDCM) where illumination is restricted to thousands of small confocal volumes during image acquisition rather than through the whole sample thus reducing photobleaching and phototoxicity.
- To follow the movement of vegetative cells frames should be captured every 2-30 sec and for developed cells every 1-20 sec. The fastest realistic frame rate is 1/sec. It may be necessary to reduce the area scanned from the default 512 pixels x 512 pixels to 512 x 300 as this allows slower scans (better images) and maximises the frame rate (Frigault et al., 2009; Muller-Taubenberger and Ishikawa-Ankerhold, 2011).
- For cells expressing the promoter_TSPOON-GFP construct typical settings used had the 488 nm laser power set at 5% and the pinhole opened up to 2.5 Airy units with a 512 pixels x 300 pixels frame collected every second. For faster temporal resolution switch to a SDCM.
- With fixed cells and ICC photobleaching can be a problem but the use of an antifade mounting medium (typically Fluoromount-G®) can minimise this and allows slower scan speeds and averaging to give better images.
- Live cell imaging is normally a balance between detecting the tagged fusion protein (signal to noise ratio) and damaging the cells with the laser due to free radicals produced by the illumination interacting with cellular constituents or the fluorophore tag. With a 10x or 20x lens this is never normally a problem but at these magnifications there is little or no spatial resolution within a cell to be gained given that Dictyostelium cells are ~10-15 µm in diameter.
- TIRF microscopy.
- The problem of phototoxicity is usually more acute with vegetative Dictyostelium cells, in part because although the evanescent field is limited to the initial ~100 nm of the sample from the coverslip, the plasma membrane is strongly illuminated.
- Image cells that have been in LOFLO or KK2C for at least one hour. The inclusion of L-Ascorbic acid may be necessary.
- Keep the laser power and the illumination time to a minimum.
- Since there is no scanning the frame rate is higher. A 100x lens (high NA) should be used with an additional 1.5x zoom lens if present as this allows ameobae to be imaged in great detail.
- Filter all buffers (0.22 µm) to remove particulates.
- Typical settings for cells expressing promoter_TSPOON-GFP construct had the 488 nm laser power set at 7% with an exposure time of 0.08 sec collecting 512 x 512 frames at up to 12 per sec.
- The problem of phototoxicity is usually more acute with vegetative Dictyostelium cells, in part because although the evanescent field is limited to the initial ~100 nm of the sample from the coverslip, the plasma membrane is strongly illuminated.
- Fixed and live cells were analysed my microscopy at the vegetative or the aggregation competent stage of development.
- Cell Fractionation
- Cells expressing A15_GFP (control) or promoter_TSPOON-GFP were grown until they reached a density of 2-4 x 106/ml in selective media, and by microscopy >50% of cells were expressing GFP.
- Starting with a maximum of 8 x 108 cells, the cells were washed in KK2 buffer and then pelleted at 300 x g for 3 min. All subsequent steps were performed at 4 °C.
- The cells were resuspended in PBS with a protease inhibitor cocktail, lysed by 8 strokes of a motorized Potter–Elvehjem homogenizer (grinding chamber clearance 0.1-0.15 mm) followed by 5 strokes through a 21-g needle to ensure full lysis.
- Alternatively, the cells can be resuspended at 1-5 x 108/ml and placed in a syringe fitted with a 25 mm filter holder containing a prefilter and a Nucleopore filter (3.0 µm). The cell suspension is then passed through this filter assembly twice.
- The lysate was then centrifuged at 4,100 x g for 32 min to pellet nuclei and unbroken cells, and the postnuclear supernatant further centrifuged at 50,000 rpm (135,700 x g RCFmax) for 30 min in a TLA-110 rotor to recover the membrane pellet and cytosolic supernatant.
- A standard protein assay was used to assess protein recovery in the 2 fractions, and volumes adjusted for equal protein.
- Alternatively, to equalise volumes, the cytosolic supernatant was concentrated by precipitation with 10% tricholoroacetic acid at 4 °C for 30 min and recovered by centrifugation 14,000 x g for 10 min. Samples precipitated with trichloroacetic acid were washed with ice cold acetone (-20 °C), air dried for 2 min and then resuspended in the same volume as the pellet fractions.
- Cells expressing A15_GFP (control) or promoter_TSPOON-GFP were grown until they reached a density of 2-4 x 106/ml in selective media, and by microscopy >50% of cells were expressing GFP.
- Protein pulldowns
Immunoprecipitations were performed using amoebae stably expressing TSPOON-GFP under a constitutive (A15_ TSPOON-GFP) and its own promoter (prom_TSPOON-GFP), and non-transformed cells were used as a control. The amounts given here are recommended for a large scale immunoprecipitation sufficient for proteomic identification of interacting proteins under native conditions. The size of the starting culture, and all subsequent volumes can be reduced accordingly for smaller scale immunoprecipitations, for instance where the identification of proteins are made by Western blotting. The protocol can also be amended for denatured (by heat and/or the presence of SDS but are not detailed here) immunoprecipitations.- Cells were grown until they reached a density of 2-4 x 106/ml in selective media, and by microscopy >50% of cells were expressing GFP.
- Up to 8 x 108 cells were pelleted by centrifugation at 300 x g for 2 min, washed twice in 50 ml of KK2 buffer before being resuspended at 2 x 107 cells/ml in KK2 buffer and starved for 4-6 h at 22 °C whilst shaking at 180 rpm.
- The cells were then pelleted at 300 x g for 3 min, lysed in 4 ml PBS-T plus protease inhibitor cocktail tablet, extracted for 20 min with rotation at 8 rpm in a 15 ml tube at 4 °C, and then spun 20,000 x g for 15 min to remove debris and insoluble material.
- By BCA protein assay the resulting lysate contained 10-15 mg total protein.
- The lysates were precleared by adding 100 µl of PA sepharose bead slurry (50% v/v in PBS) and incubated for 30 min with rotation at 8 rpm, followed by centrifugation at 2,200 x g for 3 min to pellet beads.
- The supernatant was transferred (~ 5 ml) to a fresh tube, and this is the starting material for the immunoprecipitation (protein concentration should be 2-4 mg/ml).
- For maximum recovery the lysates were immunoprecipitated using an in-house antibody against GFP overnight with rotation at 4 °C, though it may be sufficient to incubate for as little as 90 min.
- The appropriate antibody concentration requires individual optimization, though the recommended starting point is generally 2 to 5-fold higher than used for Western blotting.
- Following incubation with anti-GFP, 50 µl PA sepharose bead slurry (50% v/v in PBS) was added for 90 min at 4 °C with rotation at 8 rpm.
- An alternative would be to use a commercial source of polyclonal anti-GFP, or an anti-GFP that is already coupled to sepharose, for example GFP-TRAP that has the benefit of the antibody remaining coupled to the PA sepharose when immunoprecipitated proteins are eluted from the beads. In this case the addition of PA sepharose at this step is omitted.
- The antibody complexes recovered by centrifugation 2,200 x g for 3 min, and then washed twice with 10 ml PBS-T, resuspended in 1 ml of PBS, and transferred into a 1.5 ml microfuge tube.
- The antibody complexes were washed a further two times with PBS, pelleting the beads at 8,000 x g for 20 sec, and then eluted from the beads with 200 µl elution buffer warmed to 60 °C for 10 min.
- The beads are then pelleted at 8,000 x g for 20 sec, and the supernatant carefully removed; this can be facilitated by using a fine gel loading tip to remove the last 50 µl as the beads do not enter the tip, or by the use a micro bio-spin chromatography column.
- Eluted proteins were then precipitated with 1.2 ml acetone overnight (3-20 h) at -20 °C and centrifuged at 10,000 x g for 5 min, 4 °C. An overnight precipitation increases the yield particularly for low protein concentrations but extending it beyond this results in a lower recovery.
- The supernatant is removed and the pellet air dried for 2 min and resuspended in a buffer of choice (e.g. 1x LDS sample buffer for SDS-PAGE).
- For Western blotting of samples SDS-PAGE gels were run and transferred according to a standard protocol (Sambrook and Russel, 2001).
- For proteomics, the samples were run on pre-cast NuPAGE 4-12% Bis-Tris gels, stained with Coomassie G-250 SimplyBlue SafeStain and then cut into 8 gel slices. Each gel slice was processed by filter-aided sample preparation solution digest, and the sample was analyzed by liquid chromatography–tandem mass spectrometry in an Orbitrap mass spectrometer.
- Proteins that came down in the non-transformed control were eliminated, as were any proteins with less than 5 identified peptides, proteins that did not consistently coimmunoprecipitate in three independent experiments, or proteins of very low abundance compared with the bait (i.e., molar ratios of <0.002). The remaining proteins were considered to be specifically immunoprecipitated.
- Cells were grown until they reached a density of 2-4 x 106/ml in selective media, and by microscopy >50% of cells were expressing GFP.
Recipes
- Axenic medium
35.5 g HL5 with glucose
200 mg dihydrostreptomycin
Note: Do not use streptomycin as it is inactivated during autoclaving.
Add ultrapure dH2O to 1,000 ml
Dispense into conical flasks
75 ml/250 ml flask stoppered with 50 mm x 38 mm foam bung
150 ml/500 ml flask stoppered with 50 mm x 38 mm foam bung
750 ml/ 2,000 ml flask stoppered with 50 mm x 50 mm foam bung
The top of each flask should be covered over with a small piece (12 cm x 12 cm) of aluminium foil to prevent the top of the stopper being contaminated by dust during storage
Autoclave to sterilise
Note: 121 °C for no longer than 15 min and remove from the autoclave as soon as possible to avoid caramelisation i.e. the media turning dark brown in colour. With larger more sophisticated autoclaves with extra safety features that extend the cycle time then this may have to be reduced to as low as 7 min.
Alternatively filter sterilise (Steritop 1,000 ml unit, 0.22 µm) into sterile 1,000 ml Duran bottles or use a 1,000 ml Corning filter unit (0.22 µm)
Stored in the dark at 22 °C - LOFLO medium
8.4 g LOFLO medium
100 mg dihydrostreptomycin
Add ultrapure dH2O to 500 ml
Filter sterilise (0.22 µm) using a 500 ml Stericup filter unit
Store in the dark at 22 °C (short term) or 4 °C (long term) - SM agar
41.7 g SM agar
Add ultrapure dH2O to 1,000 ml
Autoclave to sterilise
Note: 121 °C for no longer than 15 min and remove from the autoclave as soon as possible to avoid caramelisation. With larger more sophisticated autoclaves with extra safety features that extend the cycle time then this may have to be reduced to as low as 7 minutes.
Cool to 60 °C in a heated water bath prior to pouring 25-30 ml per 10 cm diameter plastic petridish
Once the agar has set and cooled place the dishes in an airtight box or seal up in batches (15-20 plates) in plastic bags (use the bags the dishes were packed in) to minimise desiccation
Stored in the dark at 22 °C (short term <1 week) or 4 °C (long term) - SM broth
24.7 g SM broth
Add ultrapure dH2O to 1,000 ml
Dispense 10 ml into each 20 ml Universal bottle
Autoclave to sterilise
Note: 121 °C for no longer than 15 min and remove from the autoclave as soon as possible to avoid caramelisation. With larger more sophisticated autoclaves with extra safety features that extend the cycle time then this may have to be reduced to as low as 7 min.
Stored in the dark at 22 °C - KK2 buffer
2.24 g KH2PO4
0.52 g K2HPO4, anhydrous
Add ultrapure dH2O to 1,000 ml
Autoclave to sterilise in 1,000 ml Duran bottles (121 °C for 15 min)
pH after autoclaving should be ~6.1
Add 2 ml of sterile 1 M MgSO4
Stored at 22 °C - KK2C buffer
1,000 ml of sterile KK2
0.1 ml of sterile 1 M CaCl2
Stored at 22 °C - 1 M MgSO4
246.48 g MgSO4
Add ultrapure dH2O to 1,000 ml
Autoclave to sterilise in 150 ml Duran bottles (121 °C for 15 min)
Stored at 22 °C - 1 M CaCl2
147.02 g CaCl2
Add ultrapure dH2O to 1,000 ml
Autoclave to sterilise in 150 ml Duran bottles (121 °C for 15 min)
Stored at 22 °C - Dense suspension of Klebsiella aerogenes
Prepare a SM agar plate with Klebsiella streaked out on it so that there are individual colonies (stored at 8 °C and renew every 2 months from frozen stocks)
Pick a colony with a flame sterilised platinum loop (or a disposable sterile plastic needle or plastic loop) and use this to seed a universal bottle containing SM broth
Incubate the bottle for 48 h at 22 °C and vortex before using
Stored in the dark at 8 °C
Renew every month - Electroporation buffer E50
2.38 g HEPES
1.86 g KCl
0.29 g NaCl
0.5 ml of 1 M MgSO4
0.21 g NaHCO3
0.08 g NaH2PO4.2dH2O
Add ultrapure dH2O to 500 ml
Adjust to pH 7.0 with KOH
Filter sterilise (0.22 µm) using a 500 ml Stericup filter unit and stored at 4 °C - Tris-HCl buffer
0.12 g Trizma® base
Add ultrapure dH2O to 100 ml
Adjust to pH 8.0 with HCl
Autoclave to sterilise in a 150 ml Duran bottle
Stored at 22 °C - TE buffer
0.12 g Trizma® base
0.04 g EDTA
Add ultrapure dH2O to 100 ml
Adjust to pH 8.0 with HCl
Autoclave to sterilise in a 150 ml Duran bottle
Stored at 22 °C - 20 mg/ml G418
552 mg Geneticin® G418 sulphate adjusted for a potency of 724 µg/mg
Note: Potency varies from batch to batch.
Add ultrapure dH2O to 20 ml
Filter sterilise with 20 ml syringe fitted with a 0.22 µm syringe filter
Stored at -20 °C in 1 ml aliquots in sterile 1.5 ml microfuge tubes the G418 solution retains its potency for >2 years. Working stocks should be stored at 4oC and used within 6 months - Freezing medium
150 ml horse serum
11.25 ml [7.5% (v/v)] DMSO
Filter sterilise (0.22 µm) using a 150 ml Stericup filter unit
Aliquot into sterile 50 ml tubes and stored at -20 °C and should be used within 1 year - 50 mM L-Ascorbic
0.22 g L-Ascorbic
Add ultrapure dH2O to 25 ml
Filter with 50 ml syringe fitted with a 0.22 µm syringe filter to remove particulates
Stored in 0.5 ml aliquots at -20 °C - Master stock of cAMP
6.6 g cAMP
Add ultrapure dH2O to 95 ml
Adjust to pH 7.0 very carefully with KOH (Note: All the cAMP will not dissolve until the pH <5)
Add ultrapure dH2O to final volume of 100 ml
Filter sterilise (0.22 µm) using a 150 ml Stericup filter unit and stored in 20 ml aliquots at -20 °C - PBS
8.01 g NaCl
0.20 g KCl
1.42 g Na2HPO4, anhydrous
0.24 g KH2PO4
Add ultrapure dH2O to 1,000 ml
Autoclave in 500 ml Duran bottles to sterilise (121 °C for 15 min)
Stored at 22 °C - PBS-T
PBS adjusted to 1% Triton X-100, using a 10% Triton stock solution
Filter sterilise for long term storage (0.22 µm)
Stored at 22 °C or 4 °C (long term storage) - Trichloroacetic acid
100 g trichloroacetic acid
Dissolve in 35 ml ultrapure dH2O
Then make up to 100ml with ultrapure dH2O
Stored at 22 °C - Elution buffer
1.21 g Trizma® base
Add ultrapure dH2O to 75 ml
20 ml SDS 2% (w/v) from 10% solution
Adjust to pH 8.0 with HCl
Add ultrapure dH2O to a final volume of 100 ml
Filter sterilise (0.22 µm) using a 150 ml Stericup filter unit and stored in 1 ml aliquots at
-20 °C
Acknowledgments
This study was supported by the Medical Research Council [(MC_U105115237), DT and RRK] and the The Wellcome Trust (JH) .The electroporation of Dictyostelium amoebae is based on the method of Pang, Lynes and Knecht (Pang et al., 1999).
References
- Bloomfield, G., Tanaka, Y., Skelton, J., Ivens, A. and Kay, R. R. (2008). Widespread duplications in the genomes of laboratory stocks of Dictyostelium discoideum. Genome Biol 9(4): R75.
- DictyBase 1 http://www.dictybase.org/techniques/geneex/indirect_immunofluo.html.
- DictyBase 2 http://www.dictybase.org/techniques/microscopy/fixation_rgomer.html.
- Faix, J., Kreppel, L., Shaulsky, G., Schleicher, M. and Kimmel, A. R. (2004). A rapid and efficient method to generate multiple gene disruptions in Dictyostelium discoideum using a single selectable marker and the Cre-loxP system. Nucleic Acids Res 32(19): e143.
- Hagedorn, M., Neuhaus, E. M. and Soldati, T. (2006). Optimized fixation and immunofluorescence staining methods for Dictyostelium cells. Methods Mol Biol 346: 327-338.
- Hirst, J., Schlacht, A., Norcott, J. P., Traynor, D., Bloomfield, G., Antrobus, R., Kay, R. R., Dacks, J. B. and Robinson, M. S. (2014). Characterization of TSET, an ancient and widespread membrane trafficking complex. Elife 3: e02866.
- Jungbluth, A., von Arnim, V., Biegelmann, E., Humbel, B., Schweiger, A. and Gerisch, G. (1994). Strong increase in the tyrosine phosphorylation of actin upon inhibition of oxidative phosphorylation: correlation with reversible rearrangements in the actin skeleton of Dictyostelium cells. J Cell Sci 107 ( Pt 1): 117-125.
- Kimmel, A. R. and Faix, J. (2006). Generation of multiple knockout mutants using the Cre-loxP system. Methods Mol Biol 346: 187-199.
- Muller-Taubenberger, A. (2006). Application of fluorescent protein tags as reporters in live-cell imaging studies. Methods Mol Biol 346: 229-246.
- Muller-Taubenberger, A. and Ishikawa-Ankerhold, H. C. (2013). Fluorescent reporters and methods to analyze fluorescent signals. Methods Mol Biol 983: 93-112.
- Muller-Taubenberger, A., Kortholt, A. and Eichinger, L. (2013). Simple system--substantial share: the use of Dictyostelium in cell biology and molecular medicine. Eur J Cell Biol 92(2): 45-53.
- Pang, K. M., Lynes, M. A. and Knecht, D. A. (1999). Variables controlling the expression level of exogenous genes in Dictyostelium. Plasmid 41(3): 187-197.
- Sambrook, J. and Russel, D. W. (2001). Molecular cloning: A laboratory manual. Third edition. Cold Spring Harbor.
Article Information
Copyright
![]() Hirst et al. This article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0).
Hirst et al. This article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0).
How to cite
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Hirst, J., Kay, R. R. and Traynor, D. (2015). Dictyostelium Cultivation, Transfection, Microscopy and Fractionation. Bio-protocol 5(11): e1485. DOI: 10.21769/BioProtoc.1485.
- Hirst, J., Schlacht, A., Norcott, J. P., Traynor, D., Bloomfield, G., Antrobus, R., Kay, R. R., Dacks, J. B. and Robinson, M. S. (2014). Characterization of TSET, an ancient and widespread membrane trafficking complex. Elife 3: e02866.
Category
Cell Biology > Cell imaging > Fixed-cell imaging
Cell Biology > Cell imaging > Live-cell imaging
Cell Biology > Cell isolation and culture > Cell growth
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.