- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Preparation of Arabinogalactan Glycoproteins from Plant Tissue

Published: Vol 3, Iss 19, Oct 5, 2013 DOI: 10.21769/BioProtoc.918 Views: 12152
Reviewed by: Tie Liu
Original research article
The authors used this protocol in:

Jan 2013
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics







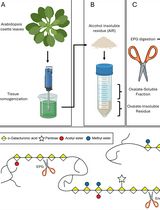
Abstract
This supplements an earlier protocol (Popper, 2011) for the extraction and assay of cell surface arabinogalactan proteins (AGPs). These highly glycosylated glycoproteins (~95% carbohydrate) contain numerous glycomodules with paired glucuronic acid residues that bind Ca2+ in a pH dependent manner (Lamport and Varnai, 2013). Classical AGPs comprise the bulk of cell surface glycoproteins and are thus integral components of a Ca2+ oscillator involved in a signalling pathway where calcium is a “universal signalling currency” analogous to ATP as the universal energy currency. The central role of these peripheral glycoproteins is thus reason enough for their further study. However, problems arise due to the extensive glycosylation and its apparent microheterogeneity generally assumed to preclude a simple reductionist approach.
Here I describe a simple partial purification of classical AGPs based on their specific interaction with the β-D-glucosyl or galactosyl Yariv reagent, a synthetic diazo dye that precipitates AGPs as an insoluble complex in salt solutions at neutral pH. (The solubility of this complex in dilute alkali provides a rapid sensitive quantitative assay for AGPs.) Reduction of the Yariv diazo linkage releases soluble AGPs for further analysis. For example deglycosylation of AGPs in anhydrous hydrogen fluoride followed by column chromatography yields just a few major AGP polypeptides purified to homogeneity (Zhao et al., 2002). However, purification of individual AGP glycoproteins to homogeneity is rarely achieved (Darjania et al., 2002); not only do the closely related AGP glycosylation profiles vastly outweigh any contribution from the amino acid composition but the glycan polydispersity made isolation of a single molecular entity well-nigh impossible until AGPs genetically engineered with a hydrophobic green fluorescent protein tag allowed chromatographic purification (Zhao et al., 2002). New approaches to AGP fractionation into discrete classes is now also a distinct possibility based on their calcium content hitherto ignored!
[Principle] Disrupted plant tissues release soluble AGPs that can be precipitated as their Yariv complex. This procedure yields mainly classical AGPs; these comprise the bulk of cell surface AGPs. Extraction with CaCl2 rather than the more usual NaCl has two advantages:
- It results in Ca2+ tightly bound by the glucuronic acid residues (Lamport and Varnai, 2013) at > pH 4.5 thus enhancing AGP solubility after its release from the insoluble Yariv complex.
- It removes pectin as insoluble calcium pectate crosslinked by intermolecular Ca2+ bridges while AGPs with intramolecular Ca2+ remain soluble.
Materials and Reagents
- Tobacco BY-2 cells or other plant tissue
- Liquid nitrogen
- CaCl2 (2% w/v)
- Distilled water
- Na metabisulphite (Sigma-Aldrich, catalog number: S-1256 )
- Dialysis tubing 12 kDa MW cutoff (3.2 cm flat width) (Sigma-Aldrich, catalog number: D-0530 )
- Superose-6, 10/300 GL (GE Healthcare, catalog number: 17-5172-01 )
- Hydroxyproline
- Glucuronic acid
- Gum arabic (Sigma-Aldrich, catalog number: G-9752 )
- NaCl
- NaOH
- Yariv reagent (Biosupplies Australia Pty, catalog number: 100-2 ) (see Recipes)
Equipment
- Blender/coffee mill
- Minifuge centrifuge
- 2 ml Sarstedt tube (with screw cap)
- Spectrophotometer or microplate reader
- Mass spectroscope
- Microfuge
- Fine-tip pipette
- Fine tip sonic probe
- Block heater
Procedure
- Freeze ~10 to 100 g plant tissue in liquid nitrogen.
- Pulverise frozen fresh tissue to a fine powder in a cold blender/coffee mill.
- Stir tissue in 2% w/v CaCl2 for 2-3 h at RT (2 ml for each gram of tissue).
- Centrifuge 30 minutes at ~10,000 x g (e.g. minifuge at RT)。
- Assay 10, 50 and 100 μl aliquots to estimate total AGP in extract (see Notes).
- Add a slight excess (see Notes) of Yariv reagent to the remaining extract.
- Allow to precipitate at least 1 h or overnight at RT.
- Collect precipitate by low speed centrifugation (10 min at 2,000 x g).
- Resuspend precipitate in 1.5 ml distilled water.
- Transfer to 2 ml Sarstedt tube (with screw cap).
- Add ~25 mg Na metabisulphite (final conc. 70 mM) to reduce the diazo linkage (top up with H2O to exlude oxygen which otherwise results in the formation of elemental sulfur).
- Cap tube tightly and heat at ~50 °C until decolourised (5-20 min).
- Transfer to small (~5 ml) dialysis bag; stir overnight at RT in 500 ml distilled H2O and change H2O three times.
- Freeze dialysate in liquid nitrogen, lyophilise, then and weigh the product. (AGP yields vary from 30-300 μg AGP/g fresh weight depending on tissue source) (see Notes).
- Validate classical AGPs by size and composition:
- Gel filtration on Superose-6 (Lamport, et al., 2006).
- Hydroxyproline content (Kivirikko and Liesmaa, 1959).
- Uronic acid content (Blumenkrantz and Asboe-Hansen, 1973).
- Bound calcium via colourimetry (Gindler and King, 1972) or ICPMS (inductively coupled plasma mass spectroscopy) (Lamport and Varnai, 2013).
- Amino acid and sugar analyses.
- Gel filtration on Superose-6 (Lamport, et al., 2006).
Notes
- Arabinogalactan protein assay via Yariv reagent.
All steps at RT.- Add test samples to Eppendorf microfuge tubes.
- Make up to ~500 μl in 1% CaCl2.
- Use 10 and 20 μg gum arabic as AGP "standards" as follows:
Add 10 μl gum arabic (1 mg/ml) to 500 μl 1% CaCl2
Add 20 μl gum arabic (1 mg/ml) to 500 μl 1% CaCl2 - Use 500 μl 1% CaCl2 as a reagent blank.
- Then to each tube:
Add 200 μl β-D-Galactosyl-Yariv reagent (1 mg/ml in 2% CaCl2)
Or use β-D-Glucosyl-Yariv reagent – choice depends on availability. - Mix well and leave for at least 30 min at room temperature.
- Spin 10 min at ~15,000 x g in microfuge.
- With CARE use fine-tip pipette to remove & discard supernate.
- Wash pellet twice with 1 ml 2% CaCl2.
- Add 1 ml 20 mM NaOH.
- Shake vigorously to dissolve pellet or sonicate 10-20 sec with fine tip probe or in a sonic bath. If solution is turbid, spin at ~15,000 x g to clarify.
- Read A457 nm against reagent blank within an hour or so. A457 avoids phenolic interference when eluting Yariv from intact BY-2 cells; for general assays read at 440 nm.
- Plot "standard curve" and calculate unknowns as μg AGP/tube.
Note: Gum arabic quantification is only an approximation as each AGP binds different amounts of Yariv.
As a general guide, however, a given weight of Yariv reagent will precipitate the same weight of AGP. So for AGP isolation a 10% excess of Yariv reagent generally suffices to precipitate all the AGP.
- Add test samples to Eppendorf microfuge tubes.
- AGP cellular distribution (background information):
Classical AGPs are essential glycoproteins distributed in three cell surface compartments: bound to the outer surface of the plasma membrane by a GPI anchor; soluble in the periplasm; and "bound" or trapped in the wall matrix.
Thus AGP cellular distribution T = M + S + W (Lamport et al., 2006)
M = AGPs bound to plasma membrane
S = Soluble AGPs released by cell disruption
W = AGPs bound to cell wall
Table 1. Total of AGPs in tobacco BY-2 cells. In tobacco BY-2 cells, T = 600 μg AGPs g fresh weight.
* AGPs upregulated by high salt appear in the growth medium.BY-2 cells(data in 6)T Total M Membrane bound S Soluble periplasmic W Wall boundSalt-adapted* 600 60 354 186 Control 600 210 282 108
Recipes
- Preparation of Yariv reagent (1,3,5-tri-(p-β-D-galactosyloxyphenylazo)-2,4,6-trihydroxybenzene) or the β-D-glucosyl derivative
Dissolve 100 mg β-D-galactosyl Yariv in 100 ml 2% w/v CaCl2
Acknowledgments
This protocol is adapted from Popper (2011), Lamport and Varnai (2013) and Lamport et al. (2006).
References
- Blumenkrantz, N. and Asboe-Hansen, G. (1973). New method for quantitative determination of uronic acids. Anal Biochem 54(2): 484-489.
- Darjania, L., Ichise, N., Ichikawa, S., Okamoto, T., Okuyama, H. and Thompson Jr, G. A. (2002). Dynamic turnover of arabinogalactan proteins in cultured Arabidopsis cells. Plant Physiol Biochem 40(1): 69-79.
- Gindler, E. and King, J. (1972). Rapid colorimetric determination of calcium in biologic fluids with methylthymol blue. Am J Clin Pathol 58(4):376-82.
- Kivirikko, K. I. and Liesmaa, M. A. (1959). A Colorimetric method for determination of hydroxyproline in tissue hydrolysates. Scandinavian J Clin Lab 11(2):128-133.
- Lamport, D. T. and Varnai, P. (2013). Periplasmic arabinogalactan glycoproteins act as a calcium capacitor that regulates plant growth and development. New Phytol 197(1): 58-64.
- Lamport, D. T., Kieliszewski, M. J. and Showalter, A. M. (2006). Salt stress upregulates periplasmic arabinogalactan proteins: using salt stress to analyse AGP function. New Phytol 169(3): 479-492.
- Popper, Z. A. (2011). Extraction and Detection of Arabinogalactan Proteins in The Plant Cell Wall - Methods and Protocols, edited by John M. Walker. Humana Press, New York, pp.245-254.
- Qi, W., Fong, C. and Lamport, D. T. (1991). Gum arabic glycoprotein is a twisted hairy rope: a new model based on o-galactosylhydroxyproline as the polysaccharide attachment site. Plant Physiol 96(3): 848-855.
- Zhao, Z. D., Tan, L., Showalter, A. M., Lamport, D. T. and Kieliszewski, M. J. (2002). Tomato LeAGP-1 arabinogalactan-protein purified from transgenic tobacco corroborates the Hyp contiguity hypothesis. Plant J 31(4): 431-444.
Article Information
Copyright
© 2013 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Lamport, D. T. A. (2013). Preparation of Arabinogalactan Glycoproteins from Plant Tissue. Bio-protocol 3(19): e918. DOI: 10.21769/BioProtoc.918.
Category
Plant Science > Plant biochemistry > Carbohydrate
Biochemistry > Protein > Isolation and purification
Biochemistry > Carbohydrate > Glycoprotein
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.