- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Conjugation Assay for Testing CRISPR-Cas Anti-plasmid Immunity in Staphylococci

Published: Vol 7, Iss 9, May 5, 2017 DOI: 10.21769/BioProtoc.2293 Views: 13185
Reviewed by: Lionel SchiavolinBenoit ChassaingDaan C. Swarts
Original research article
The authors used this protocol in:

Jan 2014
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols

Abstract
CRISPR-Cas is a prokaryotic adaptive immune system that prevents uptake of mobile genetic elements such as bacteriophages and plasmids. Plasmid transfer between bacteria is of particular clinical concern due to increasing amounts of antibiotic resistant pathogens found in humans as a result of transfer of resistance plasmids within and between species. Testing the ability of CRISPR-Cas systems to block plasmid transfer in various conditions or with CRISPR-Cas mutants provides key insights into the functionality and mechanisms of CRISPR-Cas as well as how antibiotic resistance spreads within bacterial communities. Here, we describe a method for quantifying the impact of CRISPR-Cas on the efficiency of plasmid transfer by conjugation. While this method is presented in Staphylococcus species, it could be more broadly used for any conjugative prokaryote.
Keywords: ConjugationBackground
CRISPR-Cas (Clustered, regularly interspaced, short palindromic repeats-CRISPR associated) is a prokaryotic adaptive immune system found in almost 90% of sequenced archaea and about 40% of bacteria (Makarova et al., 2015). These systems recognize and destroy nucleic acid invaders in a sequence-specific manner (van der Oost et al., 2014). A CRISPR locus typically contains an array of short DNA repeats (~35 nucleotides in length) separated by equally-short unique sequences called spacers, which are often derived from mobile genetic elements. The repeats and spacers are transcribed and processed into small CRISPR RNAs (crRNAs) that each specifies a single target. crRNAs assemble with one or more Cas proteins to form an effector complex that recognizes and degrades nucleic acids that bear a sequence, called a protospacer, complementary to the crRNA. Depending on the CRISPR-Cas type present in an organism, other limitations exist on target recognition, such as the presence of a short protospacer-adjacent motif (PAM) required for targeting by Type I and II systems (Mojica et al., 2009) or the requirement in Type III systems for specific base-pair mismatches between the crRNA and the target (Marraffini and Sontheimer, 2010). Nucleic acid invaders targeted by CRISPR-Cas are typically bacteriophages or mobile genetic elements such as plasmids or transposons that often carry virulence factors, including antibiotic resistance genes and toxins (Novick, 2003; Liu et al., 2016). CRISPR-Cas is thus a potential major barrier to the spread of virulence factors within prokaryotes and has been shown to prevent conjugative transfer of antibiotic resistance plasmids within human clinical isolates of Staphylococcus species (Marraffini and Sontheimer, 2008). Some staphylococci, including human pathogenic isolates of S. epidermidis and S. aureus, the two Staphylococcus species most commonly found in human infections, carry CRISPR-Cas systems (Cao et al., 2016; Li et al., 2016). Within these strains, multiple CRISPR spacers have been found that naturally bear homology to mobile staphylococcal plasmids, indicating that these organisms are capable of using CRISPR-Cas to limit the spread of virulence factors (Samai et al., 2015; Li et al., 2016). Investigations of the effect of mutations within the cas genes and the repeat-spacer array in Staphylococcus on anti-plasmid immunity have provided key insights into the function and mechanisms of Type III-A CRISPR-Cas in S. epidermidis RP62a (Hatoum-Aslan et al., 2011 and 2014; Maniv et al., 2016). Notably, similar assays have been used for mechanistic CRISPR-Cas studies in other organisms, including Enterococcus faecalis, Escherichia coli, and Listeria monocytogenes (Richter et al., 2014; Sesto et al., 2014; Price et al., 2016). Other methods of quantifying CRISPR anti-plasmid immunity, namely transformation via electroporation, have been used (Cao et al., 2016); however, this method cannot be used in non-competent or weakly/selectively competent organisms such as L. monocytogenes and many strains of Staphylococcus (Monk et al., 2012; Sesto et al., 2014). In these organisms, conjugation assays provide not only a viable alternative, but also a more physiologically relevant means of testing CRISPR-Cas anti-plasmid immunity. Described below is a quantitative method to determine the efficacy of CRISPR-mediated interference of the transfer of conjugative plasmid pG0400 between S. aureus and S. epidermidis. While this protocol focuses on staphylococci, it could be adapted for any prokaryotes capable of conjugation.
Materials and Reagents
- 15 ml centrifuge tubes (VWR, catalog number: 21008-216 )
- 1.7 ml microcentrifuge tubes (VWR, catalog number: 87003-294 )
- 0.45 μm membrane filters, 25 mm (EMD Millipore, catalog number: HAWP02500 )
- 50 ml centrifuge tubes (VWR, catalog number: 21008-242 )
- Pipette tips with filter, 100-1,000 μl (VWR, catalog number: 89003-060 )
- Pipette tips with filter, 1-200 μl (VWR, catalog number: 89003-056 )
- Pipette tips with filter, 1-40 μl (VWR, catalog number: 89003-048 )
- 100 x 15 mm Petri dishes (VWR, catalog number: 25384-088 )
- 0.2 ml PCR strip tubes (VWR, catalog number: 20170-004 )
- 1 mm path length cuvettes (VWR, catalog number: 97000-586 )
- Recipient S. epidermidis RP62a (Christensen et al., 1985), bearing a CRISPR-Cas system (see Note 1) (ATCC, catalog number: 35984 )
- Donor S. aureus RN4220 (Kreiswirth et al., 1983), bearing the conjugative plasmid pG0400 (Morton et al., 1995) (see Note 2) (ATCC, BEI Resources, catalog number: NR-45913 )
- Negative control S. epidermidis LAM104 (Marraffini and Sontheimer, 2008), lacking the CRISPR repeat-spacer array (see Note 3)
- Mupirocin (The United States Pharmacopeial Convention, catalog number: 1448901 )
- Brain-heart infusion broth (BD, BBL, catalog number: 211060 )
- Brain-heart infusion agar (BD, Difco, catalog number: 241830 )
- Tryptic soy broth (BD, BactoTM, catalog number: 211822 )
- Neomycin sulfate (AMRESCO, catalog number: 0558-25G )
- Brain-heart infusion broth (see Recipes)
- Brain-heart infusion agar (see Recipes)
- Tryptic soy broth (see Recipes)
Equipment
- Micropipettes set (Eppendorf, model: Research® Plus )
- 37 °C incubator (Thermo Fisher Scientific, Thermo ScientificTM, model: HerathermTM Advanced Protocol Microbiological Incubators )
- 37 °C incubated shaker capable of 160 rpm (Eppendorf, model: I26 )
- Autoclave (Getinge)
- Spectrophotometer (GE Healthcare, model: UltroSpec 10 )
- Microcentrifuge (Thermo Fisher Scientific, Thermo ScientificTM, model: HeraeusTM PicoTM 17 )
- Digital vortex mixer (VWR, model: Advanced Heavy-Duty Vortex Mixer )
- Inoculating wire loop
- Forceps
- Magnetic stir plate
Procedure
- Day 1. Preparing strains
- Streak a brain-heart infusion (BHI) agar plate containing 15 μg/ml neomycin with S. epidermidis RP62a (from here called ‘recipient’) from a -80 °C freezer stock to obtain single colonies. Incubate overnight at 37 °C.
Note: In the same manner, streak out the negative control strain, S. epidermidis LAM104. - Inoculate S. aureus carrying pG0400 (here onward called ‘donor’) from a freezer stock into 2 ml of tryptic soy broth (TSB) containing 5 μg/ml mupirocin. Incubate with shaking at 37 °C overnight.
Note: While both S. epidermidis and S. aureus strains may be propagated in either BHI or TSB, the media indicated for each strain in this protocol are consistent with the growth media used in the original protocol demonstrating anti-plasmid immunity by the Type III-A CRISPR-Cas system in S. epidermidis RP62a (Marraffini and Sontheimer, 2008). - Day 2. Growing colonies and filter mating
- After at least 16 h of incubation, large (> 1 mm in diameter) individual recipient colonies should be seen. Inoculate the entirety of 3 separate colonies into 15-ml centrifuge tubes containing 2 ml of BHI with 15 μg/ml neomycin.
Notes: - Also, inoculate any controls included in the experiment in triplicate.
- At the same time, add 50 μl of the overnight donor culture to 5 ml of TSB with 5 μg/ml mupirocin in a 15-ml centrifuge tube.
- Grow both the donor and recipient at 37 °C with shaking.
Note: At the same time, place one BHI plate per 3 growing recipient cultures into the 30 °C incubator to pre-warm (see Note 4). - After ~6 h, check the OD600 of both the recipient and donor strains.
- Add 900 μl of BHI broth into a spectrophotometer cuvette. Add 100 μl of growing culture of one of the recipient cultures. Mix well.
- After blanking with BHI, measure and record the OD600 of this 1:10 dilution of the growing culture.
- Repeat steps 2c i and 2c ii with the remaining recipient cultures.
- Repeat steps 2c i and 2c ii for the donor culture, substituting TSB for BHI.
- OD600 of all culture dilutions needs to be greater than 0.20 before proceeding. If the OD600 are not yet high enough, return to the shaker and let grow a while before checking the OD600 again.
- Calculate the amount of donor and recipient to combine to achieve a 1:4 ratio (recipients:donors) for filter mating, as in the example in Table 1.
Note: Divide 200 by the measured OD600 found in step 2c for each culture. At an OD600 of 1, there are approximately 8 x 108 cfu/ml of the donor and 2 x 108 cfu/ml of the recipient. This calculation allows for standard numbers of donors (approximately 1.6 x 109 cfu) and recipients (approximately 4 x 108 cfu) to be combined in all samples that are tested.
Table 1. Sample calculations to determine volumes of cultures to combine for filter matingCulture OD600 (1:10 dilution) 200/OD600 (Volume to add, μl) RP62a (1) 0.42 476 RP62a (2) 0.42 476 RP62a (3) 0.41 488 LAM104 (1) 0.44 455 LAM104 (2) 0.39 513 LAM104 (3) 0.44 455 RN4220/pG0400 0.56 357 - Into a 1.7 ml microcentrifuge tube, combine the above-calculated volume of the first, undiluted, recipient culture with the calculated volume of the undiluted donor.
Note: Repeat until all donor-recipient pairs are combined into separate microcentrifuge tubes. - Wash the combined cultures to remove the antibiotics by centrifuging the microcentrifuge tubes at 6,200 x g for 2 min in a table-top centrifuge. Pipette off the supernatant and resuspend the pellets in 1 ml of BHI without antibiotics. Repeat the centrifugation.
- Using sterile forceps, place one 0.45 μm membrane filter for each donor-recipient pair onto a BHI plate containing no antibiotics, with up to 3 filters on a single plate (Figure 1).
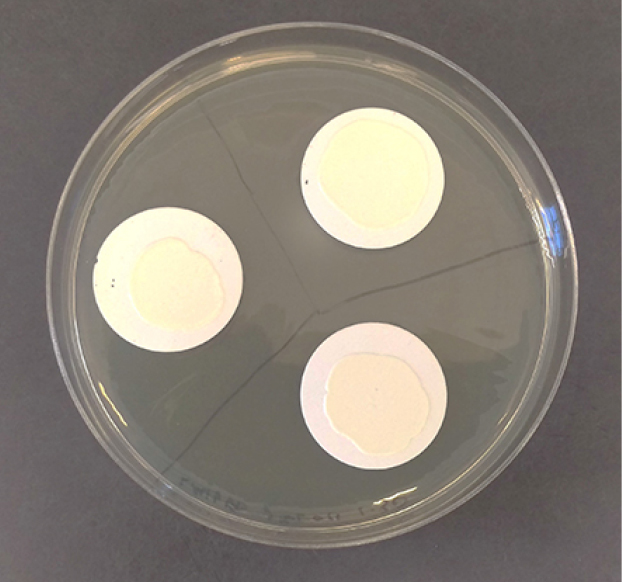
Figure 1. Sample plate for filter mating showing a plate with three membrane filters covered in overnight growth in the center of each filter - Remove the suspension from each microcentrifuge tube containing the donor and recipient combination. Resuspend the pellet in 100 μl of BHI broth and carefully pipet it onto the center of the membrane filter, being careful to not let any of the suspension fall off the membrane, as in Figure 1. It is crucial to keep all of the bacterial suspension on the filter, as this assay is quantitative and requires total recovery of all the cells.
- Repeat this with each microcentrifuge tube, placing the resuspension of each culture onto its own filter.
- Cover the plates and leave on the benchtop until the cell suspension has fully dried.
- Incubate the plates upside-down in a 30 °C incubator overnight.
- Day 3. Serially diluting and plating recipients and transconjugants
- Warm one BHI agar plate containing neomycin (15 μg/ml) and one BHI agar plate containing both neomycin (15 μg/ml) and mupirocin (5 μg/ml) for each filter from the previous night.
- In 50-ml centrifuge tubes (one for each filter), add 3 ml of BHI broth without antibiotics.
- Using sterile forceps, pick each filter off the plate and place it into a 50-ml tube prepared in step 3b.
- Vortex each tube until the cells on the filter have been fully resuspended and no growth is visible on the filter.
- Serially dilute the cell suspension
- Into each of the 8 tubes in a strip of PCR tubes, add 90 μl BHI broth.
- Remove 10 μl of the bacterial suspension from step 3d and add it to the first PCR strip tube, pipetting up and down repeatedly to resuspend (this is the 10-1 dilution).
- Using a new pipette tip, remove 10 μl from the 10-1 dilution and add it to the second PCR tube, pipetting up and down repeatedly to resuspend (this is the 10-2 dilution).
- Repeat step 3e iii to create dilutions all the way out to 10-8.
Note: Alternative methods of performing a series of 10-fold serial dilutions, such as using a multichannel pipette in a 96-well plate may be used instead. - Spot 10 μl of each dilution onto a pre-warmed BHI plate containing neomycin and another containing neomycin and mupirocin (see Note 5). All dilutions to be plated for recipient colony counting should be plated on the same plate, as in Figure 2A, and all dilutions to be plated for transconjugant colony counting should be plated on an additional plate, as in Figure 2B.
Note: Tilt the plate at a 45° angle until the drop of the dilution almost reaches the edge of the plate, then set it flat on the benchtop and let dry.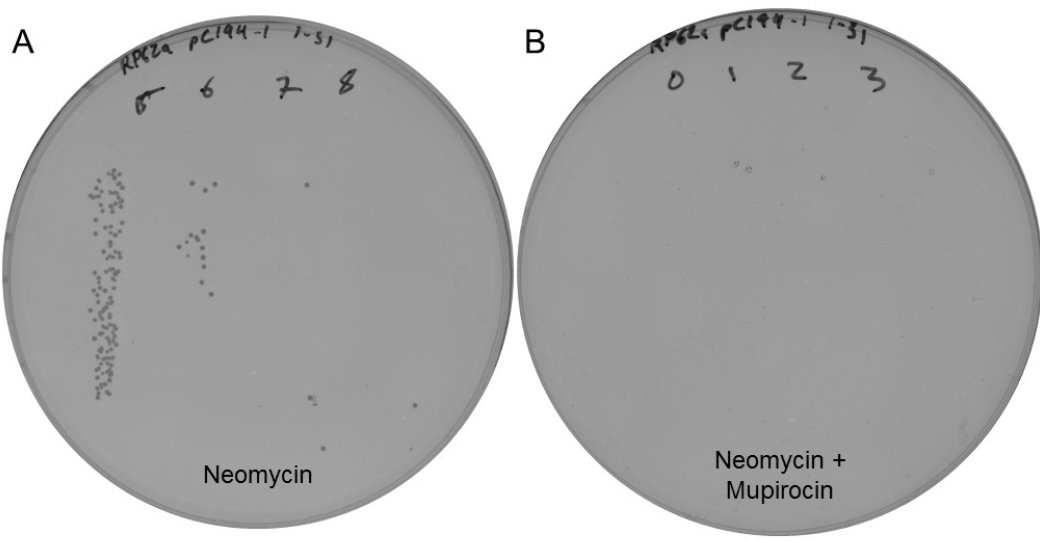
Figure 2. Sample plates from a conjugation assay using a strain of RP62a, showing recipients (A. 1.5 x 109 cfu/ml) and transconjugants (B. 0 cfu/ml) - Repeat steps 3e and 3f for each filter.
- Let the plates dry while covered on the benchtop, then incubate at 37 °C overnight.
- Day 4. Recipient and transconjugant counting
- Count the number of recipients (colonies on plates containing only neomycin) for each filter from Day 2, as shown in Figure 2A.
- Determine the number of colonies in the highest dilution where the colonies are still countable.
- Multiply the number of colonies by 100 and by the dilution factor (101 for the 10-1 dilution, 102 for the 10-2 dilution, etc.) to obtain the number of colony forming units per milliliter.
- Count the number of transconjugants (colonies on plates containing neomycin and mupirocin) for each filter from Day 2, as in step 4a and shown in Figure 2B.
- Calculate the conjugation efficiency (see Data analysis).
Data analysis
- Calculation of conjugation efficiency
- Conjugation efficiency is a measure of the number of transconjugants produced per each initial recipient cell. It can be simply calculated as follows, with an example shown in Table 2:
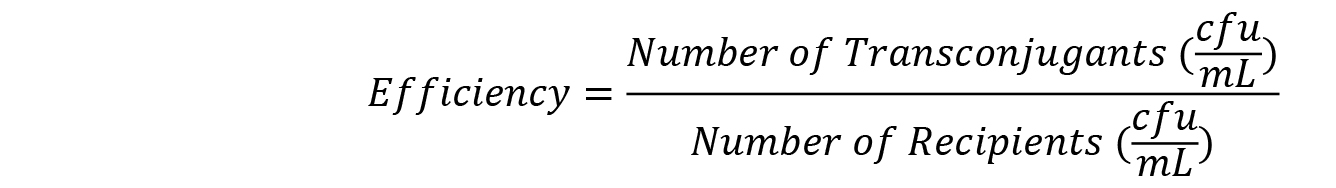
Table 2. Calculation of conjugation efficiency of a sample conjugation assayStrain Transconjugants (cfu/ml) Recipients (cfu/ml) Conjugation Efficiency RP62a (1) 1 x 103 7 x 109 1 x 10-7 RP62a (2) 2 x 102 1 x 109 2 x 10-7 RP62a (3) 0 4 x 109 0 LAM104 (1) 1.3 x 105 3 x 109 4 x 10-5 LAM104 (2) 9 x 104 3 x 109 3 x 10-5 LAM104 (3) 9 x 104 4 x 109 2 x 10-5 - A high efficiency indicates an inactive CRISPR system, while a lowered efficiency indicates active CRISPR defense.
- A 100 to 1,000-fold decrease in conjugation efficiency is typically seen in the presence of a CRISPR system as compared to a strain without a CRISPR system.
- Data presentation
Data is typically shown in a bar graph showing the number of both recipients and transconjugants in cfu/ml (colony-forming units per milliliter) on a logarithmic scale, with the calculated conjugation efficiencies accompanying the graph, as in Figure 3.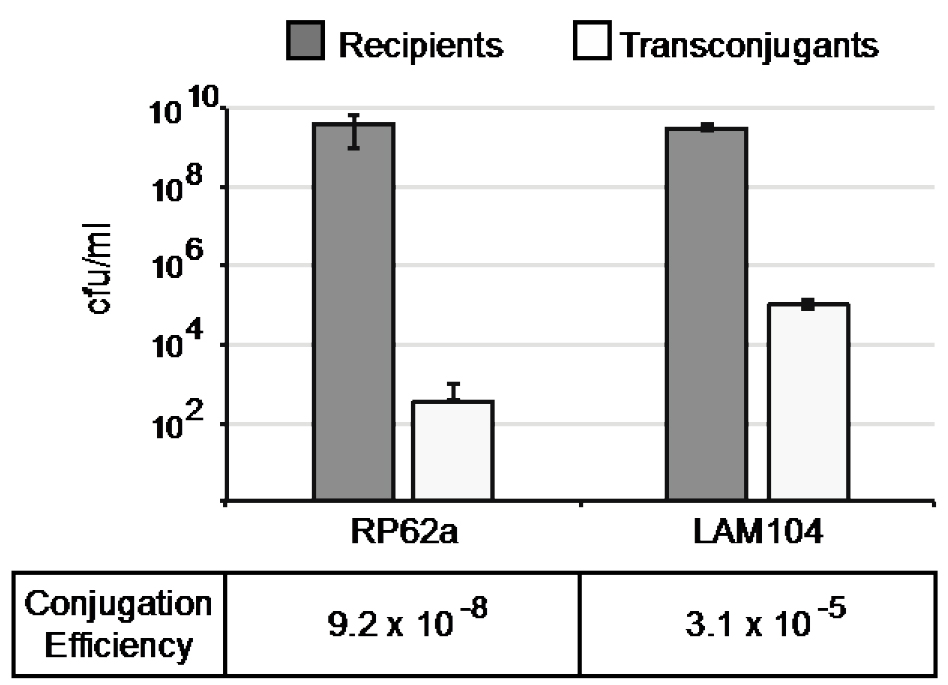
Figure 3. Sample graph of data from a conjugation assay using S. epidermidis RP62a and LAM104 as recipients. Error bars indicate the standard deviation of triplicate measurements.
Notes
- Other strains can be used as the recipient strain in this assay. S. epidermidis RP62a naturally carries a CRISPR spacer that targets the nickase gene found on conjugative staphylococcal plasmids such as pG0400 (Marraffini and Sontheimer, 2008). Other CRISPR-containing prokaryotes can be used if they are amenable to receiving conjugative plasmids. An additional important consideration in recipient selection is to make sure that the recipient has a selectable marker that differentiates it from the donor strain. In the case of conjugation assays with S. epidermidis, S. epidermidis RP62a is resistant to neomycin, so this resistance can be used to select only for transconjugants and not the donor strain itself on Day 3. Recipient strains that do not have differing antibiotic susceptibilities naturally can be transformed with a plasmid harbouring an additional resistance marker (such as pC194 [Ehrlich, 1977] for Staphylococcus). Whichever plasmid is chosen, ensure its origin is compatible with the conjugative plasmid to allow for maintenance and selection of both plasmids in the transconjugants. If using different selective markers, change antibiotics as appropriate for culturing and plating.
- Other strains of S. aureus can be used as the donor strain in this assay, as can additional conjugative plasmids. The donor strain needs to be able to perform conjugation with the recipient strain, and RN4220 is very convenient for this conjugation assay as it can perform conjugation with S. epidermidis RP62a and lacks a CRISPR system, allowing for ready uptake and maintenance of plasmids such as pG0400 that could be degraded in a CRISPR-bearing strain. Additional conjugative plasmids can also be used, depending on the CRISPR spacers found within the recipient strain. If testing to see if CRISPR functions in anti-plasmid activity in the recipient strain, a conjugative plasmid that bears a protospacer complementary to a spacer found within the CRISPR array of the recipient must be used. In the case of RP62a, pG0400 naturally has this protospacer, but in many systems, it will likely be necessary to engineer a conjugative plasmid with a protospacer. If using a different conjugative plasmid, change antibiotics as appropriate for culturing and plating. Additionally, if engineering a conjugative plasmid, be sure to follow the rules required for CRISPR targeting in the specific system to be tested, such as following the PAM requirement for Types I and II systems (Mojica et al., 2009) and the required mismatches that permit interference by Type III-A systems (Marraffini and Sontheimer, 2010).
- LAM104 is an engineered derivative of RP62a that lacks the repeat-spacer array, preventing CRISPR-Cas from functioning (Marraffini and Sontheimer, 2008). If using a strain other than RP62a, it is recommended to use a negative control in which CRISPR-Cas does not function. Suitable negative controls include 1) the use of a recipient strain that lacks a crucial component of CRISPR-Cas or 2) the use of a donor strain that carries a conjugative plasmid lacking an appropriate protospacer. S. epidermidis LAM104 is not available commercially, but can be made available from the Hatoum-Aslan laboratory on request.
- With S. epidermidis RP62a, increased background counts of recipients have been observed on Day 4 if filter mating occurs at 37 °C. To obtain a cleaner result, a 30 °C incubation is routinely used, although this can be modified to suit the growth requirements of any organism.
- With S. epidermidis strains as recipients, transconjugants are typically seen in a concentration from 100-105 cfu/ml, and therefore only the 100 to 10-3 dilutions are typically plated on the BHI plates containing neomycin and mupirocin to obtain colony counts. Recipients are usually around 109 cfu/ml and thus usually only the 10-5 to 10-8 dilutions are plated on the BHI plates containing neomycin to obtain colony counts.
Recipes
- Brain-heart infusion broth
- Dissolve 19.5 g brain-heart infusion in 500 ml deionized (DI) water
- Autoclave at 121 °C for 30 min and store at room temperature
- Brain-Heart infusion agar
- Dissolve 29 g brain-heart infusion agar in 500 ml DI water
- Autoclave at 121 °C for 30 min and stir on a magnetic stir plate until it has cooled enough to be comfortable to the touch, then add antibiotics and pour plates immediately (20 plates per 500 ml)
Note: Media can also be stored at room temperature and later heated in the microwave to liquefy and pour. - Tryptic soy broth
- Dissolve 15 g of tryptic soy broth in 500 ml DI water
- Autoclave at 121 °C for 30 min and store at room temperature
Acknowledgments
A.-H-A. is supported by the University of Alabama (UA) College of Arts and Sciences; a grant from the UA College Academy of Research, Scholarship, and Creative Activity (CARSCA); and the National Institutes of Health [5K22AI113106-02]. This protocol was adapted from that published in Marraffini and Sontheimer (2008).
References
- Cao, L., Gao, C. H., Zhu, J., Zhao, L., Wu, Q., Li, M. and Sun, B. (2016). Identification and functional study of type III-A CRISPR-Cas systems in clinical isolates of Staphylococcus aureus. Int J Med Microbiol 306(8): 686-696.
- Christensen, G. D., Simpson, W. A., Younger, J. J., Baddour, L. M., Barrett, F. F., Melton, D. M. and Beachey, E. H. (1985). Adherence of coagulase-negative staphylococci to plastic tissue culture plates: a quantitative model for the adherence of staphylococci to medical devices. J Clin Microbiol 22(6): 996-1006.
- Ehrlich, S. D. (1977). Replication and expression of plasmids from Staphylococcus aureus in Bacillus subtilis. Proc Natl Acad Sci U S A 74(4): 1680-1682.
- Hatoum-Aslan, A., Maniv, I. and Marraffini, L. A. (2011). Mature clustered, regularly interspaced, short palindromic repeats RNA (crRNA) length is measured by a ruler mechanism anchored at the precursor processing site. Proc Natl Acad Sci U S A 108(52): 21218-21222.
- Hatoum-Aslan, A., Maniv, I., Samai, P. and Marraffini, L. A. (2014). Genetic characterization of antiplasmid immunity through a type III-A CRISPR-Cas system. J Bacteriol 196(2): 310-317.
- Kreiswirth, B. N., Lofdahl, S., Betley, M. J., O'Reilly, M., Schlievert, P. M., Bergdoll, M. S. and Novick, R. P. (1983). The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature 305(5936): 709-712.
- Li, Q., Xie, X., Yin, K., Tang, Y., Zhou, X., Chen, Y., Xia, J., Hu, Y., Ingmer, H., Li, Y. and Jiao, X. (2016). Characterization of CRISPR-Cas system in clinical Staphylococcus epidermidis strains revealed its potential association with bacterial infection sites. Microbiol Res 193: 103-110.
- Liu, J., Chen, D., Peters, B. M., Li, L., Li, B., Xu, Z. and Shirliff, M. E. (2016). Staphylococcal chromosomal cassettes mec (SCCmec): A mobile genetic element in methicillin-resistant Staphylococcus aureus. Microb Pathog 101: 56-67.
- Makarova, K. S., Wolf, Y. I., Alkhnbashi, O. S., Costa, F., Shah, S. A., Saunders, S. J., Barrangou, R., Brouns, S. J., Charpentier, E., Haft, D. H., Horvath, P., Moineau, S., Mojica, F. J., Terns, R. M., Terns, M. P., White, M. F., Yakunin, A. F., Garrett, R. A., van der Oost, J., Backofen, R. and Koonin, E. V. (2015). An updated evolutionary classification of CRISPR-Cas systems. Nat Rev Microbiol 13(11): 722-736.
- Maniv, I., Jiang, W., Bikard, D. and Marraffini, L. A. (2016). Impact of different target sequences on type III CRISPR-Cas immunity. J Bacteriol 198(6): 941-950.
- Marraffini, L. A. and Sontheimer, E. J. (2008). CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA. Science 322(5909): 1843-1845.
- Marraffini, L. A. and Sontheimer, E. J. (2010). Self versus non-self discrimination during CRISPR RNA-directed immunity. Nature 463(7280): 568-571.
- Mojica, F. J., Díez-Villasenor, C., Garcia-Martinez, J. and Almendros, C. (2009). Short motif sequences determine the targets of the prokaryotic CRISPR defence system. Microbiology 155(Pt 3): 733-740.
- Monk, I. R., Shah, I. M., Xu, M., Tan, M. W. and Foster, T. J. (2012). Transforming the untransformable: application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. MBio 3(2).
- Morton, T. M., Johnston, J. L., Patterson, J. and Archer, G. L. (1995). Characterization of a conjugative staphylococcal mupirocin resistance plasmid. Antimicrob Agents Chemother 39(6): 1272-1280.
- Novick, R. P. (2003). Mobile genetic elements and bacterial toxinoses: the superantigen-encoding pathogenicity islands of Staphylococcus aureus. Plasmid 49(2): 93-105.
- Price, V. J., Huo, W., Sharifi, A. and Palmer, K. L. (2016). CRISPR-Cas and restriction-modification act additively against conjugative antibiotic resistance plasmid transfer in Enterococcus faecalis. mSphere 1(3).
- Richter, C., Dy, R. L., McKenzie, R. E., Watson, B. N., Taylor, C., Chang, J. T., McNeil, M. B., Staals, R. H. and Fineran, P. C. (2014). Priming in the Type I-F CRISPR-Cas system triggers strand-independent spacer acquisition, bi-directionally from the primed protospacer. Nucleic Acids Res 42(13): 8516-8526.
- Samai, P., Pyenson, N., Jiang, W., Goldberg, G. W., Hatoum-Aslan, A. and Marraffini, L. A. (2015). Co-transcriptional DNA and RNA cleavage during type III CRISPR-Cas immunity. Cell 161(5): 1164-1174.
- Sesto, N., Touchon, M., Andrade, J. M., Kondo, J., Rocha, E. P., Arraiano, C. M., Archambaud, C., Westhof, E., Romby, P. and Cossart, P. (2014). A PNPase dependent CRISPR system in Listeria. PLoS Genet 10(1): e1004065.
- van der Oost, J., Westra, E. R., Jackson, R. N. and Wiedenheft, B. (2014). Unravelling the structural and mechanistic basis of CRISPR-Cas systems. Nat Rev Microbiol 12(7): 479-492.
Article Information
Copyright
© 2017 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Walker, F. C. and Hatoum-Aslan, A. (2017). Conjugation Assay for Testing CRISPR-Cas Anti-plasmid Immunity in Staphylococci. Bio-protocol 7(9): e2293. DOI: 10.21769/BioProtoc.2293.
Category
Microbiology > Microbial genetics > Transformation
Microbiology > Microbial biochemistry > DNA
Molecular Biology > DNA > Transformation
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.