- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Ex vivo Culture of Fetal Mouse Gastric Epithelial Progenitors




Published: Vol 7, Iss 1, Jan 5, 2017 DOI: 10.21769/BioProtoc.2089 Views: 9888
Reviewed by: Rakesh BamHui ZhuAnonymous reviewer(s)
Original research article
The authors used this protocol in:
May 2016
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols
Abstract
Isolation and tridimensional culture of murine fetal progenitors from the digestive tract represents a new approach to study the nature and the biological characteristics of these epithelial cells that are present before the onset of the cytodifferentiation process during development. In 2013, Mustata et al. described the isolation of intestinal fetal progenitors growing as spheroids in the ex vivo culture system initially implemented by Sato et al. (2009) to grow adult intestinal stem cells. Noteworthy, fetal-derived spheroids have high self-renewal capacity making easy their indefinite maintenance in culture. Here, we report an adapted protocol for isolation and ex vivo culture and maintenance of fetal epithelial progenitors from distal pre-glandular stomach growing as gastric spheroids (Fernandez Vallone et al., 2016).
Keywords: ex vivoBackground
Mouse adult stem cells from the glandular stomach can be grown ex vivo in a 3D matrigel as ‘mini-glands’ for indefinite periods of time (Barker et al., 2010). As compared to stem cells from the small intestine growing in presence of EGF, Noggin and R-spondin 1, adult gastric stem cells need to be further supplemented with Fgf10, Gastrin, Wnt3a and a higher concentration of R-spondin 1 to get productive long-term cultures. In contrast, little was known till recently about the fetal cells that line the pre-glandular epithelium during development. So far, their nature as well as their potential growth properties ex vivo were uncharacterized. Based on the previous study identifying the cells present in the fetal small intestine (Mustata et al., 2013), we report on the culture of mouse fetal gastric progenitors as spheroids (Fernandez Vallone et al., 2016). Gastric progenitors can be replated in the culture medium previously reported by Sato et al., 2009 to grow small intestinal adult stem cells and, contrary to adult-type gastric stem cells, they do not need extra growth factors supplementation (Fgf10, Wnt3a or Gastrin).
Materials and Reagents
- Disposable scalpels (Swan Morton, catalog number: 0510 )
- Petri dishes 92 x 16 mm with cams (SARSTEDT, catalog number: 82.1473 )
- Microcentrifuge tubes, 1.5 ml (VWR, catalog number: 212-0198 )
- Tubes 10 ml, 100 x 16 mm, PP (SARSTEDT, catalog number: 62.9924.284 )
- Tubes 50 ml, 30 x 115 mm, PP (Corning, Falcon®, catalog number: 352070 )
- 70 µm nylon filters (Corning, Falcon®, catalog number: 352350 )
- P6 well plate (VWR, catalog number: 734-2323 )
- 40 µm nylon filters (Corning, Falcon®, catalog number: 352340 )
- P12 well plate (VWR, catalog number: 734-2324 )
- Tips refill (VWR, catalog numbers: 89079-464 ; 89079-470 ; 89079-478 )
- Cryotubes 1 ml (Greiner Bio One, catalog number: 123263 )
- Syringe filter 0.2 µm (VWR, catalog number: 28145-477 )
- Serological pipets 5 ml, 10 ml and 25 ml (Corning, Falcon®, catalog numbers: 357543 ; 357551 ; 357535 )
- Mice (tested on RjOrl:SWISS and C57BL/6JRj backgrounds)
- Dulbecco’s phosphate-buffered saline (DPBS), CaCl2 free, MgCl2 free (Thermo Fisher Scientific, GibcoTM, catalog number: 14190-094 )
- Fetal bovine serum (FBS) (Thermo Fisher Scientific, GibcoTM, catalog number: 10270 )
- Stem Pro Accutase cell dissociation reagent (Thermo Fisher Scientific, GibcoTM, catalog number: A1110501 )
- Matrigel® basement membrane matrix (Corning, catalog number: 354234 )
- Liquid nitrogen (supplied from Air liquide)
- Ethanol 95-97% (VWR, TechniSolv®, catalog number: 84857.360 )
- Glucose (Merck Millipore, catalog number: 1083371000 )
- Leibovitz’s L-15 medium (Thermo Fisher Scientific, catalog number: 11415-049 )
- 500 mM EDTA (pH 8.0) (Thermo Fisher Scientific, InvitrogenTM, catalog number: 15575-038 )
- Albumin from bovine serum (BSA) (Sigma-Aldrich, catalog number: A3294 )
- Advanced DMEM/F12 (Thermo Fisher Scientific, GibcoTM, catalog number: 12634-010 )
- Gentamycin 50 mg/ml (Thermo Fisher Scientific, GibcoTM, catalog number: 15750-037 )
- Penicillin-streptomycin cocktail 100x (Thermo Fisher Scientific, GibcoTM, catalog number: 15140-122 )
- Amphotericin B 250 µg/ml (Thermo Fisher Scientific, GibcoTM, catalog number: 15290-026 )
- L-glutamine (Thermo Fisher Scientific, GibcoTM, catalog number: 25030-081 )
- N-2 supplement 100x (Thermo Fisher Scientific, GibcoTM, catalog number: 17502-048 )
- B-27 w/o vit. A 50x (Thermo Fisher Scientific, GibcoTM, catalog number: 12587-010 )
- 1 M HEPES (Thermo Fisher Scientific, GibcoTM, catalog number: 15630-056 )
- N-acetyl cysteine (Sigma-Aldrich, catalog number: A7250 )
- Growth factors
Recombinant murine EGF (Peprotech, catalog number: 315-09 )
Recombinant murine Noggin (Peprotech, catalog number: 250-38 )
Recombinant murine CHO-derived R-spondin1 (R&D Systems, catalog number: 7150-RS/CF )
Rho kinase inhibitor Y27632 (Sigma-Aldrich, catalog number: Y0503 ) - DMSO (Sigma-Aldrich, catalog number: D8418 )
- Propanol-2 (VWR, catalog number: 1.09634.9900 )
- 70% ethanol (see Recipes)
- 1 M glucose (see Recipes)
- Embryo’s medium (see Recipes)
- DPBS-EDTA 5 mM (see Recipes)
- DPBS-BSA 2%-EDTA 2 mM (see Recipes)
- Basal crypt medium (BCM) (see Recipes)
- ENR medium for initial seeding (see Recipes)
- ENR medium for maintenance (see Recipes)
- Freezing medium (see Recipes)
- De-freezing medium (see Recipes)
Equipment
- Binocular (Motic, model: SMZ-168 )
- Cold light source (SCHOTT, model: KL1500 LCD )
- Scissors: straight sharp tip (Fine Science Tool, catalog numbers: 14090-09 and 14084-08 )
- Angled serrated tip forceps (Fine Science Tool, catalog number: 11080-02 )
- Standard (fine) tip forceps (Fine Science Tool, catalog number: 11251-20 )
- Micro-dissecting scissors (Fine Science Tool, catalog number: 15018-10 )
- Pipettors with Tip Ejector 20-200 µl and 100-1,000 µl (VWR, catalog numbers: 89079-970 and 89079-974 )
- Refrigerated centrifuge Refrigerated centrifuge (Beckman Coulter, model: Allegra X-15R )
- MaxQTM 4000 shaker with adaptable temperature (Thermo Fisher Scientific, Thermo ScientificTM, model: MaxQ TM 4000 )
- Biological safety cabinet (Esco Micro Pte, model: Class II Type A2 )
- Cell culture incubator (37 °C, 5% CO2) (BINDER, model: C150 )
- Inverted bright field microscope (Motic, model: AE31 )
- Nalgene Cryo ‘Mr Frosty’ freezing container (Thermo Fisher Scientific, Thermo ScientificTM, model: 5100-0050 )
- Ultra-low temperature upright freezer (Thermo Fisher Scientific, model: Thermo scientific Queue Basic )
- Cryostorage system K Series (Taylor-Wharton, model: 24K )
Procedure
Notes:
- General considerations regarding mice and pregnancies: Animals should be housed in a temperature (21 ± 1 °C) and humidity (55 ± 10%)-controlled room with a 12 h light/12 h dark cycle. Embryonic stage is calculated as embryonic day 0.5 (E0.5) the day the gestational plug is observed. It is important to remove the male from the mating cage the morning the plug is observed. Pregnancies can be followed by daily weighing of the females.
- Unless specified, steps are carried out at room temperature (RT).
- Dissection procedures and preparation of samples
- Isolation of gastric fetal epithelium for culture (Figure 1)

Figure 1. Overall procedure schematic of step A1 - General preparations before starting the dissection
Sterilize working area and dissection tools with 70% ethanol. It is not necessary to work under laminar flow, meanwhile all the steps are carried out carefully and in a clean disinfected area. It is recommended to maintain dissection tools in a glass with 70% ethanol during intermediate steps of the dissection protocol to minimize contamination.
Equipment: binocular and cold light source (Figure 2).
Dissection tools: scissors, curved forceps, standard tip forceps, scalpel, dissection bed.
Box with ice, plastic Petri dishes, embryos medium (conserved on ice), tubes with ice cold DPBS (volume according to developmental stage and number of embryos pooled – see Table 1 below and Figure 2).
Table 1. Recommended volumes (DPBS or DPBS-EDTA 5 mM or Stem Pro Accutase) for different amounts of processed tissues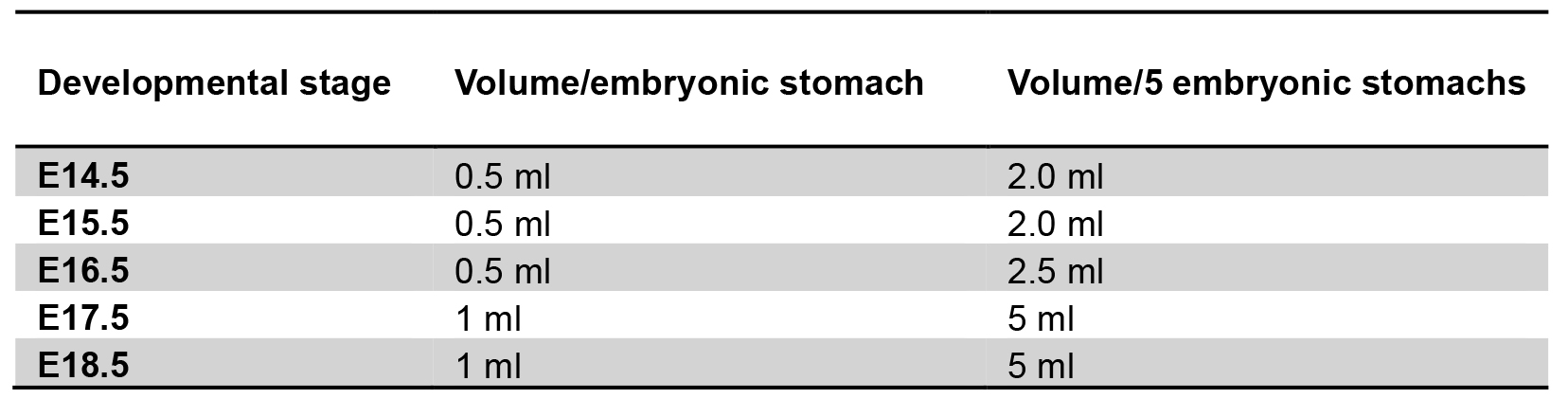

Figure 2. Example of equipment (a) and material and dissection tools (b). a. Binocular and cold light source; b. Material and dissection tools needed for embryos dissection: tubes with DPBS, Petri dish with embryos media, scalpels and straight thin tip forceps. - Euthanize the pregnant female according to the local institutional guidelines at the desired developmental stage: E14.5-E18.5. Note that visual discrimination between proximal and distal stomach areas is difficult before E14.5. Beyond E18.5 stage, spheroid yield decreases.
- Lay the pregnant female on its back and fix her to the dissection bed. Spray the abdominal area with 70% ethanol in order to sterilize the area and minimize further contamination (Figure 3a).
- Grasp the skin with angled forceps and cut transversally through the skin and peritoneum at the level of the lower abdomen enough to visualize the uterus. Uterus can be recognized as a string of pearls. Dissect out with forceps the oviduct removing the entire uterus (Figures 3b-3h).
- Place the uterus with embryos on a Petri dish filled with ice-cold embryo’s medium (see Recipes section) maintained on ice (Figure 3i).
- Separate each embryo from the uterus with the intact yolk sac using angled serrated tip forceps (Figure 3j).
- Place one of the embryos on an empty Petri dish under the binocular with cold light source. Keep the rest in the medium on ice (Figure 3k). It is recommended to perform steps A1h-l under the binocular.
- Open the yolk sac using fine straight tip forceps. Be careful not to damage the embryo’s abdomen when separating the umbilical cord (especially at E14.5-15.5 stages), as sometimes this may pull out the internal digestive tract and compromise the rest of the dissection (Figure 3l). At this point, check that the morphological embryo’s developmental stage corresponds with the one estimated by the observed vaginal plug.
- Place the embryo on its left side up and open the abdomen pulling up the skin (be careful: at early developmental stages – E14-15 the tissue is very soft and might be easily damaged). Visualize space between ribs and legs. Recognize liver-intestine-stomach block and pull it out carefully with straight thin tip forceps (Figure 3m).
- Separate the stomach from the block by ‘cleaning away’ the other organs of no interest (intestine, liver, pancreas and esophagus) using ice-cold DPBS (Figure 3n).
- To obtain higher yield of fetal spheroids, it is recommended to remove the proximal area of the stomach and continue the protocol with the distal part (Figure 3o).
Note: The elements growing in culture from the proximal part correspond to ‘squamous-like’ organoids, higher proportion of hollow fetal spheroids is obtained from the distal area (Fernandez Vallone et al., 2016).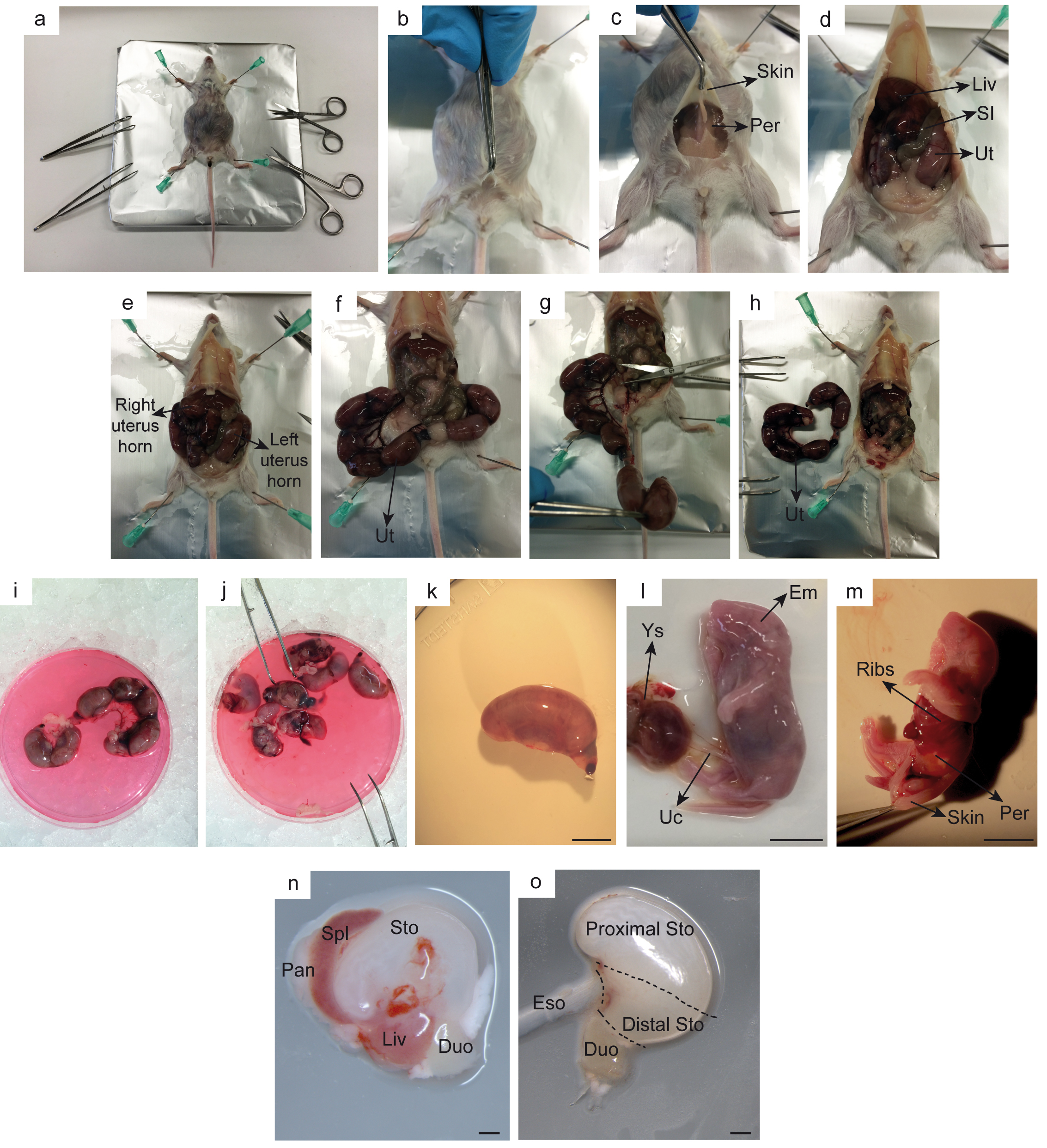
Figure 3. Representative pictures of embryonic stomach dissection at E18.5. Successive steps are detailed (a-o). Duo: duodenum, Em: embryo, Eso: esophagus, Liv: liver, Pan: pancreas, Per: peritoneum, SI: small intestine, Spl: spleen, Sto: stomach, Uc: umbilical cord, Ut: uterus, Ys: yolk sac. Scale bars: 0.5 cm (l-m); 0.1 cm (n-o). - Cut into small pieces the distal part of the stomach with the scalpel.
- Transfer the tissue to a tube containing sterile ice cold DPBS and place it on ice.
- Continue the dissection of the rest of the embryos always keeping the tissue already processed on ice.
- Centrifuge the tubes at 230 x g for 5 min, 4 °C.
- Re-suspend the pellet of each embryo or pool with the adequate volume of sterile 5 mM DPBS-EDTA (see Table 1).
- Place the tubes laid down on ice and incubate them in the shaker for 30 min with 75 rpm agitation speed.
- Centrifuge the tubes at 230 x g for 5 min, 4 °C.
- From this step onward, the protocol should be carried out under a tissue culture hood.
- Remove supernatant by aspiration and re-suspend each pellet in the adequate volume of sterile DPBS (see Table 1).
- Pipet up and down samples using FBS-coated micropipette (P1000) to disrupt the tissue and further separate the epithelium from the basal layer in contact with mesenchyme. The number of up and down pipetting times depends on the tissue developmental stage and on the number of embryos processed per sample. For example, tissues from E14-E15 individual embryos are pipetted 12 times whereas tissues from E16-E18 individual embryos or pools of embryos are pipetted between 15 and 20 times.
- Pass each suspension through a 70 µm filter into a new tube.
- Centrifuge the tubes at 300 x g for 5 min, 4 °C.
- Remove supernatant. Pellet is ready to be seeded in culture (see step B).
- Isolation of gastric fetal epithelium for culture (Figure 1)
Note: These steps allow isolating gastric fetal epithelium as clumps or group of cells that will ultimate give rise to gastric fetal spheroids in culture. However, gastric fetal spheroids can also be obtained from isolated single cells after FACS for example. In that case the user of this protocol should follow step A2 instead of step A1 (see below).
- Isolation of gastric fetal epithelium as single cells (Figure 4)

Figure 4. Overall procedure schematic of step A2- Follow protocol described in section A from step 1a to step 1o.
- Re-suspend the pellet in Stem Pro Accutase cell dissociation reagent, using volumes as described in Table 1.
- Transfer each suspension to a P6 well plate and incubate it at 37 °C with 75 rpm agitation. Time of incubation varies according to the quantity of initial material (between 30 min to 2 h). It is recommended to check the dissociation every 15 min under the inverted microscope and to help this process with mechanical up and down pipetting (micropipette P1000).
- When single cell suspension is reached, pass it through a 40 µm filter into a new tube.
- Centrifuge the tubes at 300 x g for 5 min, 4 °C.
- Remove supernatant and re-suspend the pellets in 2 ml ice cold 2 mM DPBS-BSA 2%-EDTA solution to wash.
- Repeat step A2f twice.
- Finally re-suspend the single cell preparation in 1 ml 2 mM DPBS-BSA 2%-EDTA. Maintain the cell suspension on ice and proceed with staining steps or direct sorting in case of fluorescent protein expression. It is recommended to pass the suspension through a 40 µm filter once again before sorting.
- Single cells of interest can finally be centrifuged at 300 x g for 5 min, 4 °C. Remove supernatant and if desired, proceed with sample seeding for ex vivo culture.
- Follow protocol described in section A from step 1a to step 1o.
- Ex vivo culture – 3D
- General preparations before starting
Defreeze Matrigel aliquots on ice and always keep them on ice, small changes in the temperature might accelerate undesired polymerization. - The surface of plating and amount of Matrigel will be chosen for each sample based on the size of the pellet obtained in step A. For individual samples obtained from embryos at E14-18 or sorted samples, it is recommended to use 1 well of 12 wells plate and 100 µl of Matrigel per pellet. For pooled samples obtained after step A1, 1 well of 6 wells plate and 240 µl Matrigel per pellet will be preferred. All these estimations may change according to the user’s plans and needs.
- Re-suspend the pellet in the tube with the adequate amount of Matrigel and homogenize the suspension on ice.
- Transfer the mix sample/Matrigel suspension to the plate as a drop. Stretch the drop from the center until the bottom of the well is covered without touching the walls (use a tip for this purpose) (Figure 5a).
- Place the plate in an incubator at 37 °C for 10 min until the mix polymerizes.
- Distribute the Spheroids-ENR medium for initial seeding (see Recipes section): 700 µl per well (12 well plate) and 1.4 ml per well (6 well plate).
- Place the plate in the incubator.
- Medium should be fully changed every other day with ENR medium for maintenance (see Recipes section).
Note: All steps, including the decision of plating according to density of sample should be followed by observation under inverted bright field microscope.
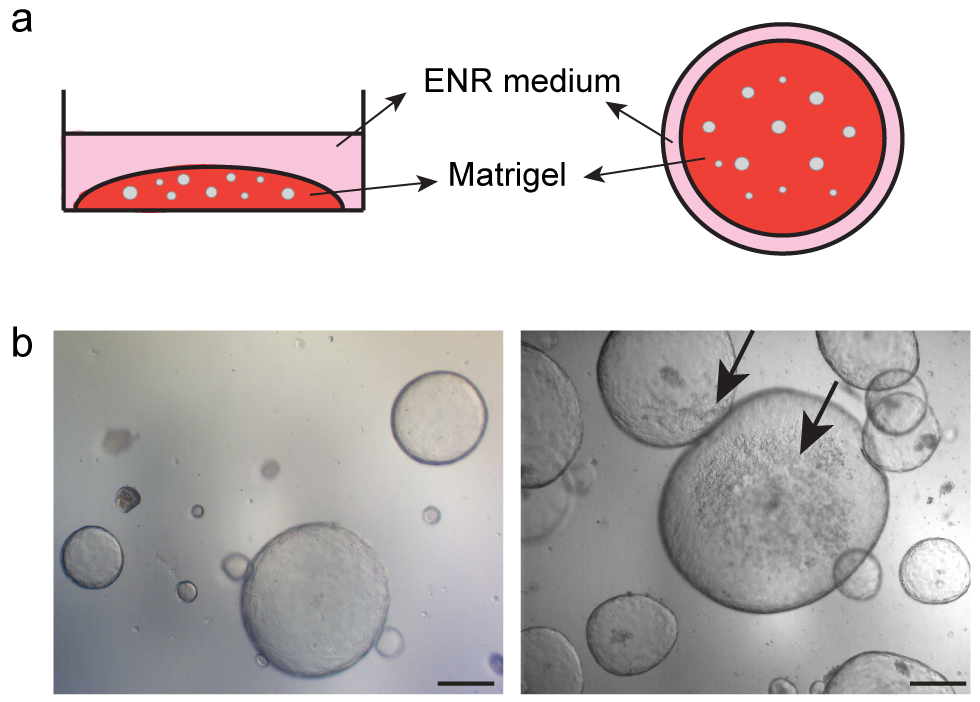
Figure 5. Spheroids initial seeding. a. Scheme showing side and upper view of the 3D culture; b. Representative pictures of gastric spheroids culture at day 6, arrows show dead cells inside the spheroids (b). Scale bars = 100 µm.
- General preparations before starting
- Maintenance of gastric spheroids
- General preparations before starting
After 5 to 7 days in culture (depending on the rate of growth), when few dead cells start to accumulate inside the spheroids, they should be replated (Figure 5b). This can be done by ‘picking’ selected elements (amplify selected population) (step C2) or by harvesting a complete part of the well (just culture maintenance) (step C3). See below details (Figure 6) for each case.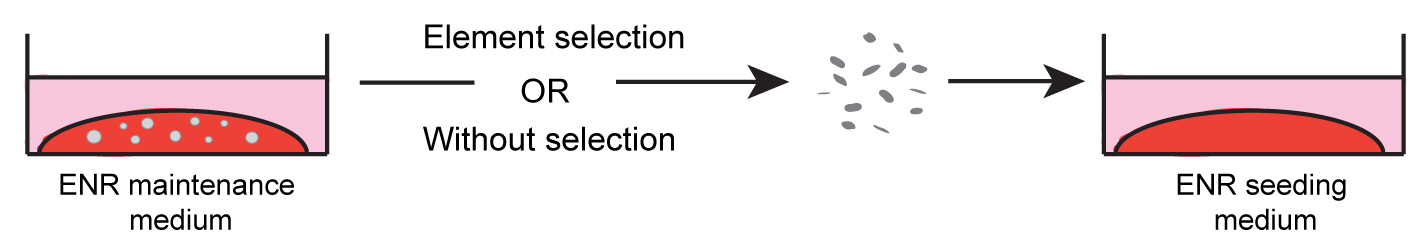
Figure 6. Procedure for maintenance of gastric spheroids. Fully grown spheroids are isolated from the Matrigel, mechanically dissociated into small groups of cells, which are then mixed with a new Matrigel aliquot and seeded. The ENR seeding medium is added after Matrigel polymerization. - Replating by element selection
- Select elements to be picked under the inverted microscope.
- Prepare one tube per sample with 1 ml DPBS at room temperature, pick the elements from the well (1 ml DPBS for 12 well plate) with a micropipette (P200) and transfer them to the tube with DPBS. Repeat the process for all the desired elements.
- Centrifuge the tube at 300 x g for 5 min.
- Remove supernatant and re-suspend the elements in 0.3 ml DPBS.
- Disrupt the elements mechanically by pipetting up and down with a micropipette (P200) until visual disappearance of big pieces.
- Add 1 ml DPBS to further wash the suspension
- Centrifuge the tube at 300 x g for 5 min.
- Remove supernatant and re-suspend the pellet in the adequate volume of Matrigel as described in step B.
- Repeat steps B3-B7.
- Select elements to be picked under the inverted microscope.
- Replating without selection
- Remove the medium from the well by aspirating.
- Add 1 ml DPBS to the well (P6 or P12 well plate) and harvest the whole Matrigel with the elements embedded by pipetting up and down with a micropipette (P1000) until homogeneous suspension is reached.
- Take 0.25 ml from the suspension (¼) and transfer it to a new tube. It is not recommended to replate the whole well unless specific need for spheroid amplification. A too high density of replating would result in too much material and debris, ending with a bad quality of replating.
- Add 1 ml DPBS to the tube and centrifuge it at 300 x g for 5 min.
- Remove supernatant and re-suspend the elements in 0.3 ml DPBS.
- Disrupt the elements mechanically by pipetting up and down with a micropipette (P200) until visual disappearance of big pieces.
- Add 1 ml DPBS to further wash the suspension.
- Centrifuge the tube at 300 x g for 5 min.
- Remove supernatant and re-suspend the pellet in the adequate volume of Matrigel as described in step B.
- Repeat steps B3-B7.
- General preparations before starting
- Cryopreservation of gastric spheroids
- Freezing protocol
- Remove the medium from the well by aspirating.
- Add 1 ml DPBS to the well and harvest the whole Matrigel with the elements embedded pipetting up and down with a micropipette (P1000) until homogeneous suspension is reached.
- Centrifuge the tube at 300 x g for 5 min.
- Wash the pellet in 2 ml DPBS.
- Centrifuge the tube at 300 x g for 5 min.
- Re-suspend the pellet in 1 ml of freezing medium (see Recipes section).
- Transfer the suspension to a labeled cryotube and place it in the Cryo freezing container (filled with isopropanol at RT). Put the container in a -80 °C freezer.
- After 48 h, cryotubes can be stored in a cryostorage system in liquid nitrogen.
- De-freezing protocol
- Warm at 37 °C a tube with 2 ml of de-freezing medium (see Recipes section).
- Take out from liquid nitrogen the selected cryotube and de-freeze the sample by pipetting up and down with pre-warmed de-freezing medium.
- Transfer all the spheroid suspension to the pre-warmed tube and centrifuge at 300 x g for 5 min.
- Remove supernatant and wash the pellet in 2 ml of BCM medium twice and centrifuge it at 300 x g for 5 min.
- Remove supernatant and re-suspend the elements in 0.3 ml DPBS.
- Disrupt the elements mechanically by pipetting up and down 20 times with a micropipette (P200).
- Add 1 ml DPBS to further wash the suspension.
- Centrifuge the tube at 300 x g for 5 min.
- Remove supernatant and re-suspend the pellet in the adequate volume of Matrigel as described in step B.
Data analysis
Details of replicates are provided in the original research paper published in free access (Fernandez Vallone et al., 2016).
Notes
- The yield of spheroid production obtained upon initial seeding is highly dependent on the developmental stage, with higher efficiency at E14-E15, and progressive decrease over time. At later developmental stages, a low proportion of clear spheroids can be obtained among the grown elements. In this case, replating of gastric spheroids will require selective picking of the clearest elements from the initial plate, the rest of the elements (usually dark spheroid-like structures) correspond to adult-type stem cells, which cannot efficiently grow in the ENR medium.
- In order to improve cell viability after cell sorting, single sorted cells are collected in the BCM medium containing 10 µM Y27632.
- This protocol can be similarly used to efficiently grow mouse fetal progenitors of the small intestine, a protocol initially described by Mustata et al. (2013).
Recipes
- 70% ethanol
70% ethanol (v/v) in distilled water - 1 M glucose
Dissolve 180 g glucose in 1 L distilled water
Pass the solution through a 0.22 µm filter - Embryo’s medium
Add 17.5 ml 1 M glucose (sterile) to 500 ml Leibovitz’s l-15 medium - DPBS-EDTA (5 mM)
0.5 ml 500 mM EDTA in DPBS
Final volume: 50 ml - DPBS-BSA 2%-EDTA (2 mM)
1 g BSA
0.1 ml 500 mM EDTA in DPBS
Final volume: 50 ml
Pass the solution through a 0.22 µm filter - Basal crypt medium (BCM)
500 ml Advanced DMEM/F12 supplemented with:
0.4 ml gentamycin
5 ml penicillin-streptomycin cocktail stock
5 ml amphotericin B
5 ml L-glutamine (final concentration: 2 mM) - ENR medium for initial seeding
BCM supplemented with:
0.5 ml N-2
1 ml B-27 w/o vit. A
0.5 ml 10 mM HEPES
0.1 ml 1 mM N-acetyl cysteine
Growth factors at a final concentration of: 50 ng/ml for EGF, 100 ng/ml for Noggin and 100 ng/ml for CHO-derived R-spondin 1, 10 µM Rho kinase inhibitor (Y-27632)
Final volume: 50 ml - ENR medium for maintenance
BCM supplemented with:
0.5 ml N-2
1 ml B-27 w/o vit. A
0.5 ml 10 mM HEPES
0.1 ml 1 mM N-acetyl cysteine
Growth factors at a final concentration of: 50 ng/ml for EGF, 100 ng/ml for Noggin and 100 ng/ml for CHO-derived R-spondin1
Final volume: 50 ml - Freezing medium
BCM
1 ml FBS
1 ml DMSO
Final volume: 10 ml - De-freezing medium
BCM
1 ml FBS
Final volume: 10 ml
Acknowledgments
This work was supported by the Interuniversity Attraction Poles Programme-Belgian State-Belgian Science Policy (6/14), the Fonds de la Recherche Scientifique Médicale of Belgium, the Walloon Region (program CIBLES) and the non-for-profit Association Recherche Biomédicale et Diagnostic. This protocol was adapted from previous work for sample preparation (Mustata et al., 2013) and the initial report of Sato et al. (2009) for ex vivo culture conditions.
References
- Barker, N., Huch, M., Kujala, P., van de Wetering, M., Snippert, H. J., van Es, J. H., Sato, T., Stange, D. E., Begthel, H., van den Born, M., Danenberg, E., van den Brink, S., Korving, J., Abo, A., Peters, P. J., Wright, N., Poulsom, R. and Clevers, H. (2010). Lgr5+ve stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro. Cell Stem Cell 6(1): 25-36.
- Fernandez Vallone, V., Leprovots, M., Strollo, S., Vasile, G., Lefort, A., Libert, F., Vassart, G. and Garcia, M. I. (2016). Trop2 marks transient gastric fetal epithelium and adult regenerating cells after epithelial damage. Development 143(9): 1452-1463.
- Mustata, R. C., Vasile, G., Fernandez-Vallone, V., Strollo, S., Lefort, A., Libert, F., Monteyne, D., Perez-Morga, D., Vassart, G. and Garcia, M. I. (2013). Identification of Lgr5-independent spheroid-generating progenitors of the mouse fetal intestinal epithelium. Cell Rep 5(2): 421-432.
- Sato, T., Vries, R. G., Snippert, H. J., van de Wetering, M., Barker, N., Stange, D. E., van Es, J. H., Abo, A., Kujala, P., Peters, P. J. and Clevers, H. (2009). Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 459(7244): 262-265.
Article Information
Copyright
© 2017 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Vallone, V. F., Leprovots, M., Vassart, G. and Garcia, M. (2017). Ex vivo Culture of Fetal Mouse Gastric Epithelial Progenitors. Bio-protocol 7(1): e2089. DOI: 10.21769/BioProtoc.2089.
Category
Stem Cell > Adult stem cell > Maintenance and differentiation
Cell Biology > Cell isolation and culture > 3D cell culture
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.