- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Determination of Rate of [3H-methyl]-choline Incorporation into Cellular Lipids and Non-lipid Metabolites
Published: Vol 6, Iss 22, Nov 20, 2016 DOI: 10.21769/BioProtoc.2019 Views: 8159
Reviewed by: Neelanjan BoseYong TengCarsten Ade
Original research article
The authors used this protocol in:

Mar 2016
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols

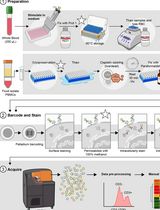
Abstract
The choline-containing phospholipid, phosphatidylcholine (PtdCho) is the most common mammalian phospholipid found in cell membrane (Ide et al., 2013). It is also a component of intracellular signalling pathways (Cui and Houweling, 2002). Herein is described a measure of the rate of accumulation of choline by lipid soluble PtdCho and lyso-Ptdcho which can further be discriminated by chromatographic analysis (Smith and Phyu, 2016). Determination of the accumulation of [3H-methyl]-choline into water-soluble components is also described. The procedure could be used to measure the effect of drugs and other factors on choline incorporation into phospholipids. After exposure of cells to test conditions (e.g., drugs) adherent cells in tissue culture flasks are incubated with radiolabelled [3H-methyl]-choline in medium for 15 min (pulse). The [3H-methyl]-choline is then rapidly removed and incubation continued in the presence of non-radioactive medium (chase). Cellular distribution of [3H-methyl] is then determined by cell fractionation and measurement of radioactivity in the lipid and non-lipid cellular components.
Background
Phospholipid metabolism is essential in formation of cell membranes (Ide et al., 2013) and cell signalling (Cui and Houweling, 2002). Both the formation of choline-containing metabolites and the accumulation of choline into lipids are pivotal processes during cellular proliferation. Perturbations in phospholipid metabolisms are associated with cancer and other disorders (Gibellini and Smith, 2010). Measurement of these processes is central to understanding medical imaging modalities that detect choline incorporation by tumour tissue using [11C]-choline-PET (positron emission tomography) (Podo et al., 2007) and choline metabolite content using 31P or 1H (proton) magnetic resonance spectroscopy (Saeedi et al., 2005).
Here is described a method of quantitating the incorporation of choline into aqueous and lipid components. The method is straightforward, relatively inexpensive and easy to set up. It is also quantitative. Alternative methods include NMR spectroscopy (Mori et al., 2015) which can be used to measure the content of phospholipids in tissues and tissue extracts but requires very expensive equipment and specialist knowledge and thin layer chromatography which is inexpensive but is considered to be a qualitative technique.
Materials and Reagents
- Reaction tubes 1.5 ml (Greiner Bio one, catalog number: 616201 )
- Scintillation vials 20 ml (PerkinElmer, catalog number: 6000477 )
- 200 µl pipette tips
- 1 ml pipette tips
- Microfuge tubes
- Trypsin/EDTA, 0.05% trypsin/0.53 mM EDTA in HBSS (w/o) calcium and magnesium (Mediatech, catalog number: 25-051-CI )
- Fetal bovine serum (FBS) (Thermo Fisher Scientific, GibcoTM, catalog number: 10270106 )
- Penicillin-streptomycin (10,000 U/ml) (Thermo Fisher Scientific, GibcoTM, catalog number: 15140122 )
- [3H-methyl]-choline (specific activity 2.22-3.33 TBq/mmol) (American Radiolabelled Chemicals, catalog number: ART 0197-1 mCi )
- Phosphate-buffered saline (PBS, 10x) diluted 10x with water (Thermo Fisher Scientific, Fisher Scientific, catalog number: 12579099 )
- Tissue culture medium Dulbecco’s modified Eagle’s medium (DMEM) with glutamax (Thermo Fisher Scientific, GibcoTM, catalog number: 31966021 )
- Methanol (Sigma-Aldrich, catalog number: 34860 )
- Chloroform (Sigma-Aldrich, catalog number: C2432 )
- Trizma-hydrochloride buffer solution pH 7.4 (Sigma-Aldrich, catalog number: T2194 )
- Scintillation fluid Ultima Gold XR (PerkinElmer, catalog number: 6013329 )
- Bicinchoninic Acid Kit (Sigma-Aldrich, catalog number: BCA1 )
- Zinc sulfate monohydrate (ZnSO4·H2O) (Sigma-Aldrich, catalog number: 96495 )
- Barium hydroxide octahydrate [Ba(OH)2·8H2O] (Sigma-Aldrich, catalog number: B2507 )
- [3H-methyl]-choline in medium (see Recipes)
- 5% ZnSO4 (w/v) (see Recipes)
- Ba(OH)2 solution (see Recipes)
- HCl (1 N) (see Recipes)
- NaOH (1 N) (see Recipes)
Equipment
- Tissue culture flasks 80 cm2 growth surface (Thermo Fisher Scientific, Thermo ScientificTM, catalog number: 178905 )
- Tissue culture flasks 25 cm2 growth surface (Thermo Fisher Scientific, Thermo ScientificTM, catalog number: 136196 )
- Incubator (CO2) (Thermo Fisher Scientific, Fisher Scientific, catalog number: 15311035 )
- Centrifuge (Thermo Fisher Scientific, Thermo ScientificTM, model: Heraeus Fresco 21 )
- Scintillation counter (PerkinElmer, model: tricarb 4910 )
- Pipettes P1000 (Gilson, catalog number: F123602 )
- Pipettes P200 (Gilson, catalog number: F132601 )
- Pipettes P20 (Gilson, catalog number: F132600 )
- Vortex mixer (Whirlimix) (Cole-Parmer Instrument, catalog number: UY-04726-01 )
Procedure
- Grow (adherent) cells until 80% confluent in 80 cm2 tissue culture flasks.
- Detach cells with a solution of 0.05% trypsin: EDTA (5 ml) and neutralise with same volume of medium containing 10% fetal bovine serum (FBS) and penicillin-streptomycin (10,000 U/100 ml).
- Add 1 ml of cell suspension containing 0.5 x 106 cells (this is for cells with doubling times of about 48 h – if the doubling time is slower or faster adjust seeding number accordingly so that the cells are 60-80% confluent on the day of measurement of [3H-methyl]-choline uptake) to each* 25 cm2 tissue culture flask required and make up to 5 ml with medium.
*Note: Typically triplicates are used for each treatment and also for control (untreated) so if an experiment compares the effect of two treatments on lipid metabolism then 9 flasks need to be seeded with cells. - Incubate flasks for 24 h at 37 °C in a CO2 with a humidified gas mix of 5% CO2:95% air incubator after which they can be treated with drugs if desired.
- After the desired drug incubation period remove medium and add 1 ml of medium containing [3H-methyl]-choline (37 KBq/ml) – see Recipe 1. Incubate for 15 min at 37 °C.
Note: Residual media in the flasks will dilute the added radioactive medium so fully remove by suction. - At the end of the incubation period with [3H-methyl]-choline rapidly add 5 ml of ice cold phosphate buffered saline (PBS) and discard by inverting the flask into a sink (designated for aqueous radioactive waste) and rinse 3 more times with ice-cold PBS (5 ml).
- The cells are then incubated for 1 h with fresh 1 ml of non-radioactive medium (chase). During this time the [3H-methyl]-choline moves through the phosphatidylcholine anabolic pathway and accumulates as phosphatidylcholine and its catabolites.
- At the end of the chase period the media is collected and the cells washed 2 x with 0.2 ml PBS and the washes pooled with the medium. Radioactivity is counted in the pooled media/washes by adding to a scintillation vial containing scintillation fluid (at a ratio of 1:10 solution:scintillation fluid) and measuring in the liquid scintillation counter. This is carried out so that the amount of activity that has been lost by efflux during the chase period can be determined if required.
- The cells are then detached by addition of 0.3 ml of a solution of 0.05% trypsin-EDTA (Gibco UK) and incubation to allow detachment to occur followed by neutralization of trypsin with 0.3 ml of medium containing 10% FBS.
- Using a P1000 pipette transfer the cell suspension to a reaction tube.
- Centrifuge the cells at 1,000 x g for 5 min at 4 °C then the media is removed and retained. During the trypsin treatment some of the cytosolic contents may leak out of the cells so if a measure of total activity in the cell is required the activity in the post-trypsinisation washes should be measured.
- The cells are then lysed by resuspension in 0.25 ml of methanol (MeOH):CHCl3 (2:1) and placing on ice for 1 h after briefly vortex mixing.
- The cell contents are fractionated into aqueous and lipid soluble phases by addition of 0.125 ml of CHCl3 and 0.125 ml of Tris buffer (10 mM) to the lysed cells and separating by centrifuging at 10,000 x g for 5 min at 4 °C.
- The aqueous (upper phase) is then collected into a separate tube (see Figure 1). Removal of the aqueous phases is aided by tilting the tube at about a 45° angle and drawing off the liquid into a 200 µl pipette tip (using a P200 Gilson pipette). The aqueous volumes are typically 0.5 ml so repeated fillings of the 200 µl pipette tip are required but the smaller tip is more controllable than using a 1 ml pipette tip.
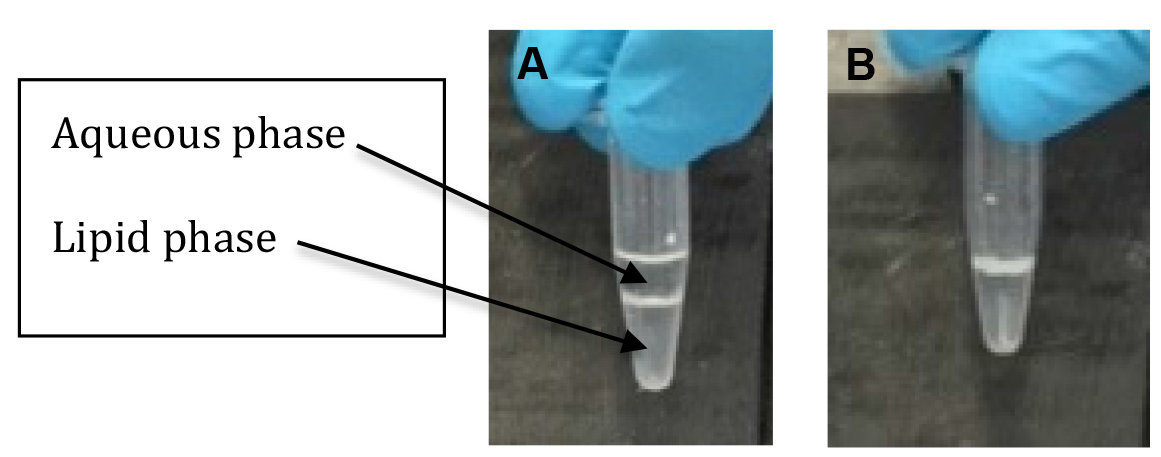
Figure 1. Appearance of the aqueous and lipid phases and precipitated protein interface after centrifugation. A. The aqueous (upper), protein interface and lipid (lower) phases after centrifugation and B. after removal of the aqueous phase. - The lipid (lower phase), containing the fraction of [3H-methyl]-choline incorporated into lipid, is removed, leaving behind the protein precipitate, placed in a scintillation vial with scintillation fluid (at a 1:10 ratio of lipid phase to scintillation fluid) mixed and the activity counted in the liquid scintillation counter. Removal of the lower phase can be tricky as it consists mainly of chloroform which, being heavy, can leak out of the pipette tip but is achieved by gently drawing up the fluid into a 200 µl pipette tip using a P200 pipette and transferring rapidly to the scintillation vial (keeping close at hand). A 200 µl tip is preferable to a 1 ml tip as it’s easier to draw up the small fiddly volumes (typically 0.3 ml).
- The residual solvents in the tube now containing just the protein precipitate is left to dry at room temperature and then the pellet is dissolved overnight in 100 μl of NaOH (1 N).
Note: Complete drying is important as the solvents may interfere with the protein assay. Drying can be speeded up by placing in an incubator at 37 °C. - After addition of 100 µl of HCl (1 N) to neutralise the NaOH, the amount of protein in the sample is then determined using a protein assay kit (e.g., Bicinchoninic Acid Kit) and following the manufacturer’s instructions.
- Discrimination of phosphorylated and non-phosphorylated [3H-methyl]-choline in the aqueous phase from step 13 (adapted from Kletzien and Perdue, 1974) (This technique can also be used to discrimination of phosphorylated and non-phosphorylated in the cell washes from step 10 or the media from step 7):
- The total volume of the aqueous phase is determined by iteratively drawing the liquid into a pipette tip on a P1000 pipette and adjusting the volume until the pipette tip is fully loaded when all the liquid has been drawn up into the pipette tip.
- Two 100 µl samples of the liquid is placed in two microfuge tubes. To one of them is added 200 μl of water and after vortex mixing 100 µl is placed in a scintillation vial containing 1 ml of scintillation fluid (1:10 ratio of liquid to scintillation fluid) and the radioactivity counted. This is the total phosphorylated + non-phosphorylated [3H-methyl]-choline in 33.3 µl of the aqueous phase.
- To the other 100 µl sample is added, each of 100 µl of ZnSO4 (5%, w/v) and 100 μl of 0.3 M Ba(OH)2. After mixing on a vortex for 5 min the precipitated phosphates (including those containing [3H-methyl]-choline phosphates) are pelleted by centrifugation at 10,000 x g for 10 min and 100 µl of supernatant is removed to a scintillation vial containing 1 ml of scintillation fluid (1:10 ratio of liquid to scintillation fluid) to determine non-phosphorylated [3H-methyl]-choline by radioactivity counting in the scintillation counter.
- The amount of [3H-methyl]-choline in 33.3 µl of aqueous phase is determined by subtracting the non-phosphorylated [3H-methyl]-choline from the total [3H-methyl]-choline from step 18b.
- The total volume of the aqueous phase is determined by iteratively drawing the liquid into a pipette tip on a P1000 pipette and adjusting the volume until the pipette tip is fully loaded when all the liquid has been drawn up into the pipette tip.
Data analysis
- Incorporation of [3H-methyl]-choline into lipid: The incorporation of [3H-methyl]-choline into lipid is then normalised for cell number by dividing the radioactive counts in the lipid fraction by the protein content (in mg).
- Incorporation of [3H-methyl]-choline into the aqueous phase: The incorporation of [3H-methyl]-choline into aqueous components is determined by correcting the radioactive counts in the aqueous phases for dilution. Steps 17a-17d determined the amount of phosphorylated and non-phosphorylated [3H-methyl]-choline in 33.3 µl of the aqueous phase. To obtain the total amounts of each in the aqueous phase multiply by the volume of the aqueous phase in µl divided by 33.3. Both the phosphorylated and non-phosphorylated components are normalized for the total protein content determined for the corresponding sample. The data can be displayed as a histogram, e.g., Figure 2.
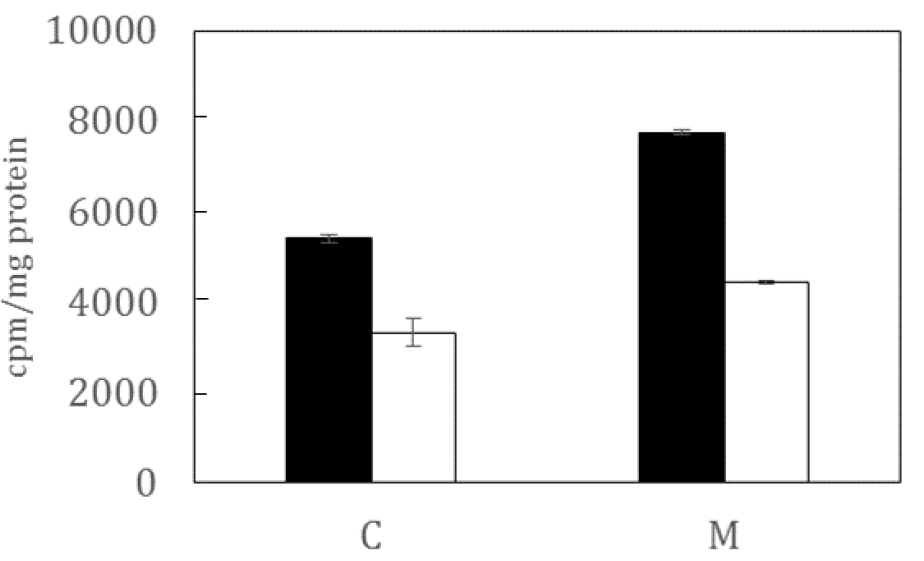
Figure 2. Representation of typical data showing [3H-methyl]-choline incorporation by untreated (C) and metformin(M)-treated tumour cells in the absence (black) and presence (white) of an inhibitor of phosphatidylinositol-phospholipase C
Note: The details of number of sample replicates, number of independent experiments and statistical tests recommended for this type of work can be found in Smith and Phyu (2016).
Recipes
- [3H-methyl]-choline in medium (make up fresh for each experiment)
Each flask will require 1 ml of medium containing 37 KBq of [3H-methyl]-choline
Using a G20 pipette add the requisite volume of [3H-methyl]-choline to the requisite volume of medium for an experiment. E.g., if the experiment involves 9 flasks, add 9 x 37 KBq of [3H-methyl]-choline to 9 ml of medium.
Notes:- Personal protective equipment (gloves and goggles) should be worn at all times when handling radioactivity.
- Pipette tips used to transfer radioactive liquids should be washed several times with water before disposal.
- Personal protective equipment (gloves and goggles) should be worn at all times when handling radioactivity.
- 5% ZnSO4 (w/v)
0.5 g ZnSO4
10 ml water - Ba(OH)2 solution (0.3 M)
1.71 g Ba(OH)2
10 ml water - HCl (1 N)
8.2 ml of concentrated (37%) HCl
100 ml water - NaOH (1 N)
4.0 g NaOH
100 ml water
Acknowledgments
The work described herein was funded by a grant from Grampian Hospital NHS Endowment. This work is modified from earlier studies by the author’s group (Al-Saeedi et al., 2005)
References
- Cui, Z. and Houweling, M. (2002). Phosphatidylcholine and cell death. Biochim Biophys Acta 1585(2-3): 87-96.
- Gibellini, F. and Smith, T. K. (2010). The kennedy pathway - De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB LIFE 62: 414-428.
- Kletzien, R. F. and Perdue, J. F. (1974). Sugar transport in chick embryo fibroblasts. I. A functional change in the plasma membrane associated with the rate of cell growth. J Biol Chem 249(11): 3366-3374.
- Ide, Y., Waki, M., Hayasaka, T., Nishio, T., Morita, Y., Tanaka, H., Sasaki, T., Koizumi, K., Matsunuma, R., Hosokawa, Y., Ogura, H., Shiiya, N. and Setou, M. (2013). Human breast cancer tissues contain abundant phosphatidylcholine(36:1) with high Stearoyl-CoA Desaturase-1 expression. PLOS ONE 8(4): e61204.
- Mori, N., Wildes, F., Kakkad, S., Jacob, D., Solaiyappan, M., Glunde, K. and Bhujwalla, Z. M. (2015). Choline kinase-α protein and phosphatidylcholine but not phosphocholine are required for breast cancer cell survival. NMR Biomed 28(12): 1697-1706.
- Podo, F., Saradanelli, F., Iorio, E., Canese, R., Carpinelli, G., Fausto, A. and Canevari, S. (2007). Abnormal choline phospholipid metabolism in breast and ovary cancer: molecular bases for noninvasive imaging approaches. Curr Med Imaging Rev 3(2): 123-137.
- Saeedi, F. A., Welch, A. E. and Smith, T. A. D. (2005). [methyl-3H]-choline incorporation into MCF7 tumour cells: correlation with proliferation. Eur. J. Nucl. Med. 32(6):660-667.
- Smith, T. A. and Phyu, S. M. (2016). Metformin decouples phospholipid metabolism in breast cancer cells. PLoS One 11(3): e0151179.
Article Information
Copyright
© 2016 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Smith, T. A. D. and Phyu, S. M. (2016). Determination of Rate of [3H-methyl]-choline Incorporation into Cellular Lipids and Non-lipid Metabolites. Bio-protocol 6(22): e2019. DOI: 10.21769/BioProtoc.2019.
- Smith, T. A. and Phyu, S. M. (2016). Metformin decouples phospholipid metabolism in breast cancer cells. PLoS One 11(3): e0151179.
Category
Biochemistry > Lipid > Lipid measurement
Cell Biology > Cell signaling > Intracellular Signaling
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.