- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Detection of HBV C Protein Phosphorylation in the Cell

Published: Vol 5, Iss 15, Aug 5, 2015 DOI: 10.21769/BioProtoc.1551 Views: 8617
Reviewed by: Smita NairVarpu MarjomakiAnonymous reviewer(s)
Original research article
The authors used this protocol in:

Aug 2014
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics








Abstract
Among the seven serines and one threonine in the carboxyl-terminus of HBV C protein, all but one (serine 183) appear in the context of RxxS/T consensus phosphoacceptor motifs and also overlap with other consensus motifs, such as S/TP, RS, SPRRR, RRRS/T, or RRxS/T, suggesting that various cellular kinases phosphorylate these residues. To determine whether threonine and/or serine (serines 157, 164, 170, 172, 178, and 180, and threonine 162, adw subtype) of HBV C protein are indeed phosphoacceptor residues in cells, Huh7 were transfected with a series of C-protein-expressing mutants, labeled with 32P-orthophosphate for 14 h, and then lysed. The 32Pi-labeled lysates were immunoprecipitated with anti-HBc antibody, and the 32Pi-labeled immunoprecipitated C proteins were detected by autoradiography.
Materials and Reagents
- Huh7 hepatoma cells (Japanese Collection of Research Bioresources Cell Bank, catalog number: JCRB0403 )
- Dulbecco’s modified Eagle’s medium (DMEM) (Life Technologies, Gibco®, catalog number: 12800-017 )
- Fetal bovine serum (FBS) (Life Technologies, Gibco®, catalog number: 16000-044 )
- Penicillin/streptomycin (Life Technologies, Gibco®, catalog number: 15140-122 )
- Plasmid; HBV WT C protein (STSSSS) in plasmid HBV-P-def of pcDNA3 backbone, phosphoacceptor-site mutants (ATAAAA, AAAAAA, SSSSSS, and ASAAAA) in plasmid HBV-P-def in pcDNA3 backbone, pcDNA3-GFP
- Polyethylenimine (Polysciences, catalog number: 23966 )
- Opti-MEM (Life Technologies, Gibco®, catalog number: 31985-062 )
- OptiMEM (Life Technologies, Gibco®, catalog number: 31985-070 )
- Dialyzed FBS (Life Technologies, Gibco®, catalog number: 26400044 )
- 1 mCi orthophosphate [32Pi] (PerkinElmer Inc., catalog number: NEX053 )
- Polyclonal rabbit anti-HBc antibody (home-made, Jung et al., 2012)
- Protein A/G Plus agarose beads (Calbiochem®, catalog number: IP05 )
- Tris-HCl (pH 8.5) (Sigma-Aldrich, catalog number: T6066 )
- EDTA (Sigma-Aldrich, catalog number: E5134 )
- Nonidet P-40 (Sigma-Aldrich, catalog number: CA630 )
- NaF (Sigma-Aldrich, catalog number: 201154 )
- β-glycerophosphate (USB, catalog number: 155-56-2 )
- Sodium orthovanadate (Sigma-Aldrich, catalog number: s6508 )
- Protease inhibitors (Calbiochem®, catalog number: 535142 )
- Tris-HCl (pH 6.8) (Sigma-Aldrich, catalog number: T6066)
- SDS (Sigma-Aldrich, catalog number: L-3771 )
- β-mercaptoethanol (Sigma-Aldrich, catalog number: M6250 )
- Bromophenol blue (Sigma-Aldrich, catalog number: 114391 )
- Glycerol (USB, catalog number: 16374 )
- Lysis buffer (see Recipes)
- 2x sample buffer (see Recipes)
Equipment
- 10-cm dishes (Corning Incorporated, catalog number: 430167 )
- PVDF membranes (Bio-Rad Laboratories, AbD Serotec®, catalog number: 162-0177 )
Procedure
- Huh7 hepatoma cells were grown in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin under a humidified atmosphere at 37 °C in 5% CO2.
- Cells were passaged every three days (passage at 80~90% confluency) and 2 x 106 Huh7 cells were seeded in 10-cm dishes one day before the transfection.
- Next day, Huh7 cells were transfected using polyethylenimine (PEI).
PEI transfection method:- In a sterile tube, total 10 μg of plasmid DNA was mixed with 500 μl Opti-MEM.
- Add 30 μl of PEI solution (1 μg/1 μl) to DNA-Opti-MEM solution and then vortex immediately.
- Incubate 15 min at room temperature.
- Then add PEI/DNA-Opti-MEM mixture to cells.
- In a sterile tube, total 10 μg of plasmid DNA was mixed with 500 μl Opti-MEM.
- Transfection experiments were repeated at least three times.
- Forty-eight hours after transfection, Huh7 cells were incubated at 37 °C in 5% CO2 incubator for 6 h in 10 ml DMEM containing 10% dialyzed FBS, then labeled at 37 °C in 5% CO2 for additional 14 h with 250 µCi orthophosphate [32Pi] per 107 cells, rinsed with ice-cold phosphate-buffered saline, and lysed with 1 ml of lysis buffer. (Isotope was added directly onto the cells containing DMEM-dFBS.)
- Lysates were transferred to 1.5 ml tube, incubated on ice for 10 min for further lysis, spin down at 13,000 rpm at 4 °C for 2 min, and supernatant was transferred to fresh 1.5 ml tube.
- The supernatant lysates were incubated for 2 h at 4 °C with 1 μl of polyclonal rabbit anti-HBc antibody on the rotator.
- Next, 15 μl of protein A/G Plus agarose beads was added, and the mixture was incubated at 4 °C for 1 h and centrifuged at 1,500 rpm for 10 sec. (Protein A/G Plus agarose bead preparation: Take 15 μl of bead/sample to fresh 1.5 ml tube, add 1 ml lysis buffer, vortex briefly, spin down at 1,500 rpm for 10 sec, and discard supernatant. Repeat above washing procedures 2 more times.)
- The pellets in 1.5 ml tube (containing beads bound to immune complexes of radiolabeled WT or mutant C proteins and anti-HBc antibodies) were washed twice with 1 ml lysis buffer, eluted by boiling (100 °C for 5 min) in 15 μl of 2x sample buffer, and separated by SDS-PAGE on 13.5% gels. Running time was 3 h at 80 voltage.
- Separated proteins were transferred to PVDF membranes (250 mA 1 h 30 min) and subjected to autoradiography.
Representative data
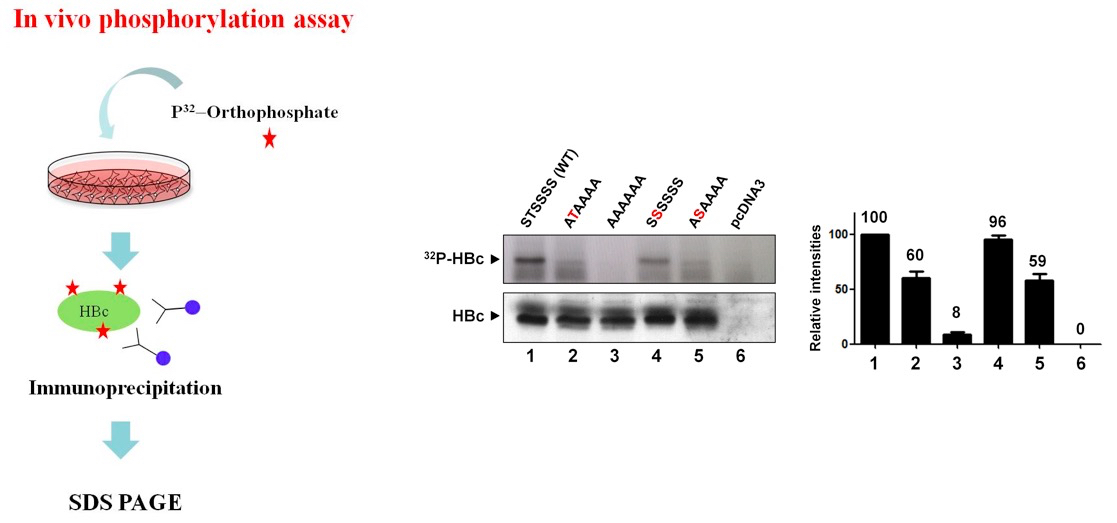
Compared to AAAAAA mutant of which the phosphoacceptor sites were all abolished to alanine, STSSSS (WT) and other mutants were all 32Pi-labeled.
Recipes
- Lysis buffer
50 mM Tris-HCl (pH 8.5)
2 mM EDTA
0.5% Nonidet P-40
50 mM NaF
25 mM β-glycerophosphate
2 mM sodium orthovanadate, and protease inhibitors - 2x sample buffer
100 mM Tris-HCl (pH 6.8)
4% SDS
200 mM β-mercaptoethanol
0.2% bromophenol blue
20% glycerol
Acknowledgments
This work was supported by National Research Foundation Grants funded by the Korean Government (NRF-2012-R1A2A2A01015370).
References
- Jung, J., Kim, H. Y., Kim, T., Shin, B. H., Park, G. S., Park, S., Chwae, Y. J., Shin, H. J. and Kim, K. (2012). C-terminal substitution of HBV core proteins with those from DHBV reveals that arginine-rich 167RRRSQSPRR175 domain is critical for HBV replication. PLoS One 7(7): e41087.
- Jung, J., Hwang, S. G., Chwae, Y. J., Park, S., Shin, H. J. and Kim, K. (2014). Phosphoacceptors threonine 162 and serines 170 and 178 within the carboxyl-terminal RRRS/T motif of the hepatitis B virus core protein make multiple contributions to hepatitis B virus replication. J Virol 88(16): 8754-8767.
Article Information
Copyright
© 2015 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Jung, J. and Kim, K. (2015). Detection of HBV C Protein Phosphorylation in the Cell. Bio-protocol 5(15): e1551. DOI: 10.21769/BioProtoc.1551.
Category
Microbiology > Microbial biochemistry > Protein > Modification
Biochemistry > Protein > Immunodetection > Western blot
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.