- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Seed Coat Ruthenium Red Staining Assay
Published: Vol 4, Iss 7, Apr 5, 2014 DOI: 10.21769/BioProtoc.1096 Views: 20195
Reviewed by: Renate WeizbauerRu ZhangAnonymous reviewer(s)
Original research article
The authors used this protocol in:

Jul 2013
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics






Related protocols

Abstract
The goal of this protocol is to assay for defects in synthesis/secretion/release of seed coat mucilage by ruthenium red staining of mature whole seeds.
The mucilage secretory cells of the Arabidopsis seed coat synthesize and secrete a large quantity of primarily pectinaceous mucilage to a ring-shaped apical domain during their differentiation. This makes them an excellent model system to identify genes involved in both cell wall synthesis and secretion (et al., 2000). When wild-type seeds are incubated in ruthenium red stain, hydrated mucilage is extruded from epidermal cells and a ‘halo’ of red-stained mucilage is observed surrounding the seed (Western et al., 2000). Reduced mucilage staining may result from defects in cell wall biosynthesis, secretion, or impaired release upon hydration.
Materials and Reagents
- Seeds
- 0.01% ruthenium red solution (Sigma-Aldrich, catalog number: 11103-72-3 )
- 50 mM EDTA (optional)
- dH2O (pH 6.5-7.5) [buffer with 10 mM Tris (pH 7.6), if necessary]
Equipment
- Microfuge tubes
- Transfer pipet or aspirator
- Orbital shaker
- White porcelin depression plate or clean depression slides
- Dissecting or compound microscope
Procedure
- Place ~20 seeds in a clean (but not necessarily sterile) microfuge tube.
- Add 800 μl dH2O or 50 mM EDTA.
Shake vigorously (~400 rpm) on an orbital shaker for 2 h at room temperature to hydrate, and therefore release the mucilage from the epidermal seed coat (see Note 6).
- Remove as much of the first solution as possible with a pipet or an aspirator and replace with 800 μl of 0.01% ruthenium red solution.
- Shake vigorously (~400 rpm) on an orbital shaker for 1 h at room temperature.
- Remove the ruthenium red solution and replace with dH2O.
- Observe the seeds, mounted in water, under a dissecting microscope in a white porcelin depression plate, or on a clean depression slide under a compound microscope.

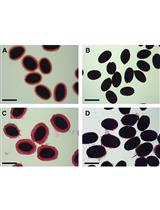
Figure 1. Mucilage staining of WT and echidna mutant seeds incubated in EDTA and ruthenium red solution (Gendre et al., 2013). Wild-type seeds hydrated and stained with ruthenium red display a ‘halo’ of stained mucilage around the seed. This halo was almost completely absent in echidna seeds indicating a putative defect in mucilage biosynthesis, secretion, or release.
Notes
- Ruthenium red is a non-specific chemical dye that binds to negative charges, therefore it will stain the acidic polysaccharides that make up pectin.
- EDTA treatment chelates Ca2+ ions from solution, thereby reducing pectin crosslinking. Thus, EDTA treatment can particularly be used to promote the release of mucilage both from mutants with reduced amounts of mucilage (e.g. Arsovski et al., 2009a) and those with chemically altered mucilage that has altered hydration and swelling (e.g. Arsovski et al., 2009b).
- If dH2O is significantly lower than pH 7, the dye won't work very well.

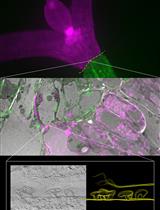
Figure 2. Effect of the pH and EDTA on the ruthenium staining. WT seeds were incubated for 2 h in water with an acidic pH 5.8, a buffered Tris solution or EDTA, prior to the ruthenium red staining. The acidity is affecting the staining intensity. The EDTA treatment promote mucilage release giving rise to a thicker halo.
- Seeds should be completely desiccated (1-2 weeks after harvesting minimum) before staining.
- Environmental variations in plant growth can lead to differences in mucilage. If plants are under stress conditions (i.e. water stress, pathogen attack, deficiency or excess of nutriments) the seed quality, and therefore mucilage production, may be affected. It is best to compare seed mucilage from plants that were grown together, and/or to repeat the experiment several times on different batches of seeds.
- Seeds have two layers of mucilage: a loose outer layer that washed away with shaking, and a more tightly bound layer that remains associated with the seed even after shaking or washing. To observe both layers, seeds can be stained without shaking in individual wells of a tissue culture plate (e.g. 24-48 well plate) (Harpaz-Saad et al., 2011).
Recipes
- 0.01% ruthenium red
0.1 g in 1 L dH2O (pH 6.5 -7.5)
Stored at 4 ºC
Discard solution if no longer bright pink (It may turn brownish, orange, purple.)
- 50 mM EDTA
1,861 g in 100 ml dH2O
Add EDTA to 80 ml of dH2O
Bring pH to ~8-9 (until EDTA dissolves) with NaOH pellets or 10 N NaOH solution (~2 g of pellets)
Bring volume to 100 ml
Acknowledgments
This protocol was developed by T. Western at the University of British Columbia while in the laboratory of Dr. George Haughn, funded through National Sciences and Engineering Research Council grants to G. Haughn. While the first use was shown in Western et al. (2000) referenced in the text, the protocol was first described in Western et al. (2001) and elaborated for details of EDTA pretreatment in Arsovski et al. (2009a), referenced in the text.
References
- Arsovski, A. A., Villota, M. M., Rowland, O., Subramaniam, R. and Western, T. L. (2009a). MUM ENHANCERS are important for seed coat mucilage production and mucilage secretory cell differentiation in Arabidopsis thaliana. J Exp Bot 60(9): 2601-2612.
- Arsovski, A. A., Popma, T. M., Haughn, G. W., Carpita, N. C., McCann, M. C. and Western, T. L. (2009b). AtBXL1 encodes a bifunctional beta-D-xylosidase/alpha-L-arabinofuranosidase required for pectic arabinan modification in Arabidopsis mucilage secretory cells. Plant Physiol 150(3): 1219-1234.
- Gendre, D., McFarlane, H. E., Johnson, E., Mouille, G., Sjodin, A., Oh, J., Levesque-Tremblay, G., Watanabe, Y., Samuels, L. and Bhalerao, R. P. (2013). Trans-Golgi network localized ECHIDNA/Ypt interacting protein complex is required for the secretion of cell wall polysaccharides in Arabidopsis. Plant Cell 25(7): 2633-2646.
- Harpaz-Saad, S., McFarlane, H. E., Xu, S., Divi, U. K., Forward, B., Western, T. L. and Kieber, J. J. (2011). Cellulose synthesis via the FEI2 RLK/SOS5 pathway and cellulose synthase 5 is required for the structure of seed coat mucilage in Arabidopsis. Plant J 68(6): 941-953.
- Western, T. L., Skinner, D. J. and Haughn, G. W. (2000). Differentiation of mucilage secretory cells of the Arabidopsis seed coat. Plant Physiol 122(2): 345-356.
- Western, T. L., Burn, J., Tan, W. L., Skinner, D. J., Martin-McCaffrey, L., Moffatt, B. A. and Haughn, G. W. (2001). Isolation and characterization of mutants defective in seed coat mucilage secretory cell development in Arabidopsis. Plant Physiol 127(3): 998-1011.
- Western, T. L. (2012). The sticky tale of seed coat mucilages: production, genetics, and role in seed germination and dispersal. Seed Sci Res 22(1): 1.
Article Information
Copyright
© 2014 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
McFarlane, H. E., Gendre, D. and Western, T. L. (2014). Seed Coat Ruthenium Red Staining Assay. Bio-protocol 4(7): e1096. DOI: 10.21769/BioProtoc.1096.
Category
Plant Science > Plant physiology > Tissue analysis
Biochemistry > Carbohydrate > Glycoprotein
Cell Biology > Tissue analysis > Tissue staining
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.