

Optimized Mechanical Isolation of Mitochondria From Saccharomyces cerevisiae Preserving Atg32 for Quantitative Analysis
保留 Atg32 的酿酒酵母线粒体优化机械分离方法用于定量分析
(§Technical contact: amendoza@ifc.unam.mx) 发布: 2026年02月20日第16卷第4期 DOI: 10.21769/BioProtoc.5610 浏览次数: 523
评审: Hemant Kumar PrajapatiPriyanka MittalAnonymous reviewer(s)
Advertisement










Abstract
Mitophagy is a highly conserved process among eukaryotic cells, playing a primordial role in mitochondrial quality control and overall cellular homeostasis. In Saccharomyces cerevisiae, Atg32 is the only identified mitophagy receptor localized to the mitochondrial outer membrane, making this yeast a particularly powerful model for molecular studies of mitophagy that require the isolation of intact mitochondria. However, traditional methods for isolating mitochondria from yeast often rely on enzymatic cell wall digestion and homogenization, which can compromise the stability of mitochondrial surface proteins such as Atg32. In this protocol, we describe an optimized mechanical approach for yeast cell disruption using glass beads in a cold, protease-inhibited buffer to preserve mitochondrial integrity and facilitate the detection of Atg32. Subsequent differential centrifugation and washing steps yield mitochondrial fractions suitable for downstream biochemical analyses. This workflow eliminates enzymatic digestion steps, reduces sample variability, and allows parallel processing of multiple strains or experimental conditions. Overall, this method offers a rapid, low-cost, and reproducible alternative for crude mitochondrial isolation, ensuring excellent preservation of Atg32 and broad compatibility with quantitative and comparative studies.
Key features
• Mechanical cell disruption using glass beads preserves mitochondrial integrity and enables reliable immunodetection of Atg32 without requiring enzymatic spheroplasting.
• Rapid, low-cost, and highly reproducible workflow suitable for processing multiple yeast strains or experimental conditions in parallel.
• Optimized cold, protease-inhibited lysis conditions minimize Atg32 degradation and improve detection sensitivity in mitochondrial fractions.
Keywords: Mitophagy (线粒体自噬)Graphical overview
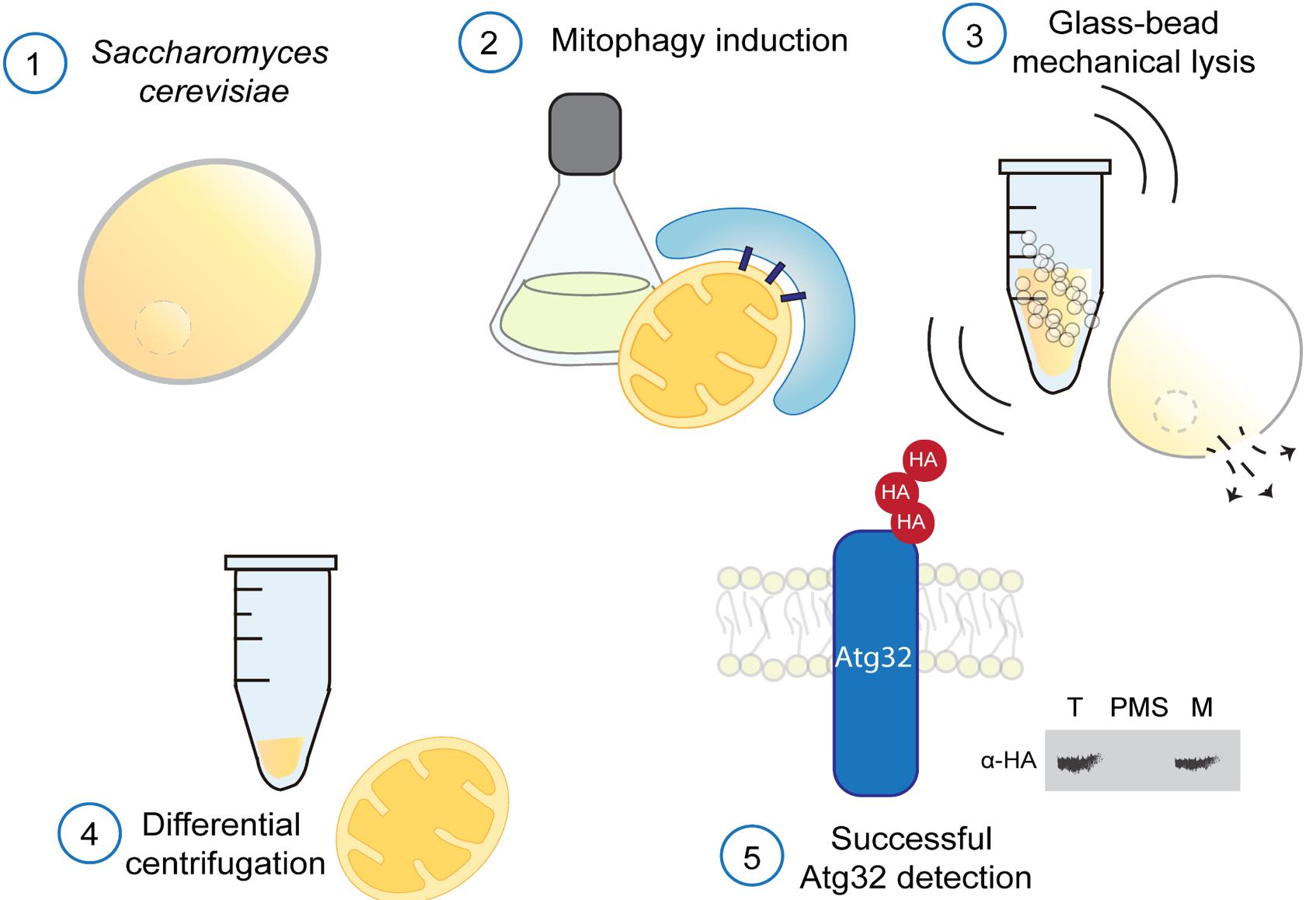
Saccharomyces cerevisiae cells subjected to mitophagy-inducing conditions are lysed by glass-bead vortexing under strictly cold conditions, with the addition of protease inhibitors to preserve mitochondrial integrity and Atg32 stability. Crude extracts are fractionated by differential centrifugation to obtain total (T), post-mitochondrial supernatant (PMS), and mitochondrial (M) fractions. 3HA-Atg32 is detected by immunoblotting.
Background
Mitochondrial quality control is essential for cellular homeostasis and is maintained through dynamic processes, including mitochondrial degradation (mitophagy) and mitochondrial biogenesis. Mitophagy is a degradation process of mitochondrial portions that depends on the autophagic machinery [1]. For mitochondria selective recognition, several receptor proteins have been identified at the outer mitochondrial membrane in metazoans [2–8]. However, the yeast Saccharomyces cerevisiae utilizes a single well-characterized receptor, Atg32, which plays a central role in initiating and regulating this process [9,10].
Atg32 is an outer mitochondrial membrane protein with cytosolic and intermembrane space–exposed domains, which acts as a molecular bridge between mitochondria and the autophagic machinery [10,11].
Given the importance of mitophagy as a conserved eukaryotic mechanism for mitochondrial turnover, it is crucial to understand the regulation of this protein under both non-induced and induced mitophagy conditions. However, reproducible biochemical approaches for in situ immunodetection of Atg32 within mitochondrial fractions are limited in the current literature [9,10,12,13]. Existing mitochondria isolation protocols typically rely on enzymatic cell wall digestion with zymolyase, followed by Dounce homogenization [14], which is labor-intensive, costly, and often leads to proteolytic degradation of Atg32. As shown in Figure 1A, the traditional zymolyase-based method results in a complete loss of detectable Atg32 in mitochondrial fractions under both non-mitophagy and mitophagy-induced conditions, severely limiting its utility for studying its regulation. Such proteolytic susceptibility renders this approach unreliable and introduces variability that compromises experimental reproducibility (Figure 1C). In contrast, using our glass bead–based mechanical lysis method, the Atg32 band is clearly detectable in mitochondrial fractions under both non-induced and mitophagy-induced conditions (Figure 1B). Although Atg32 levels are reduced in total fractions upon mitophagy induction, consistent with mitochondrial degradation [10,13,15–17], the protein remains readily detectable, enabling reliable comparative analyses (Figure 1D). Our method eliminates the need for enzymatic digestion and homogenization, resulting in a faster, more cost-effective, and highly reproducible protocol that preserves mitochondrial integrity and allows detection of Atg32.
Materials and reagents
Biological materials
For Atg32 detection, a triple haemagglutinin epitope was added to the N-terminus of Atg32 (3HA-Atg32) by fusion PCR. This version was expressed from a centromeric plasmid (pRS316) [18] under the regulation of endogenous ATG32 promoter and terminator (pRS316-3HA-ATG32 plasmid) [16]. The resulting plasmid was used to transform a Δatg32 strain.
Table 1. Yeast strains
| Strain | Genotype | Reference |
|---|---|---|
| EY0986 (IDH1-GFP) Δatg32[pRS316-3HA-ATG32] | EY0986 (IDH1-GFP) atg32::KanMX4 [pRS316-3HA-ATG32] | [16,17] |
Reagents
1. Acrylamide (Bio-Rad, catalog number: 1610140)
2. Adenine (Sigma, catalog number: A2786)
3. Ammonium persulfate (APS) (Sigma, catalog number: A3678)
4. Ammonium sulfate (Merck, catalog number: 8090260)
5. Anti-HA peroxidase high affinity (3F10) (Roche, catalog number: 12013819001)
6. Anti-Oxa1 (N-terminus); kind gift from Johannes M. Hermann’s Lab
7. Anti-Pgk1 (Thermo Fisher Scientific, catalog number: 459250; 1:5000)
8. β-mercaptoethanol (Sigma, catalog number: MG250)
9. Bromophenol blue (Amresco, catalog number: 0449)
10. BSA stock; Quick Start Bovine Gamma Globulin (BGG) Standard (Bio-Rad, catalog number: 5000208)
11. Chemiluminescent substrate (Immobilon Western, catalog number: WBKLS0100)
12. Complete Mini, EDTA-free (Roche, catalog number: 11836170001)
13. Glucose (Sigma, catalog number: G8270)
14. Glycerol (J.T. Baker, catalog number: 2136-02)
15. Glycine (Sigma, catalog number: G7126)
16. HCl (J.T. Baker, catalog number: 9535-02)
17. HEPES (Sigma, catalog number: H3375)
18. Histidine (Sigma, catalog number: H-8000)
19. L-Lysine (Sigma, catalog number: L5501)
20. L-Methionine (Sigma, catalog number: M9625)
21. Leucine (Formedium, catalog number: DOC0156)
22. Low-fat milk (Nestlé-Svelty, commercial mark)
23. N,N,N′,N′-Tetrametiletilendiamina (TEMED) (Sigma, catalog number: T9281)
24. N,N′-Methylenebisacrylamide solution (Bis-acrylamide) (Sigma, catalog number: M1533)
25. Phenylmethylsulfonyl fluoride (PMSF) (Sigma, catalog number: P-7626)
26. Protein marker ladder (NIPPON Genetics, catalog number: MWP06)
27. Quick Start Bradford 1× dye reagent (Bio-Rad, catalog number: 5000205)
28. Sodium chloride (NaCl) (J.T. Baker, catalog number: 3624-01)
29. Sodium dodecyl sulfate (SDS) (Sigma, catalog number: L3771)
30. Sorbitol (Sigma, catalog number: S1876)
31. Tris-HCl (Roche, catalog number: 1185-53-1)
32. Trizma base (Sigma, catalog number: T1503)
33. Tryptophan (Formedium, catalog number: DOC0188)
34. Tween 20 (Sigma, catalog number: P6585)
35. Yeast nitrogen base w/o amino acids and ammonium sulfate (Difco, catalog number: 233520)
Solutions
1. Synthetic minimal glucose (SMG) (see Recipes)
2. Synthetic minimal glycerol (SMGly) (see Recipes)
3. Buffer SHP (see Recipes)
4. TBS 10× (see Recipes)
5. TBS-Tween 10× (see Recipes)
6. LAE 2× (see Recipes)
7. SDS-PAGE: running gel 14% (see Recipes)
8. SDS-PAGE: stacking gel 4.85% (see Recipes)
9. Running buffer 10× (see Recipes)
Recipes
Note: All percentages are w/v unless otherwise indicated.
1. Synthetic minimal glucose (SMG)
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| Yeast nitrogen base | 1.7 g/L | 1.7 g |
| Ammonium sulfate | 5 g/L | 5 g |
| Glucose 40% (v/v) | 2% (v/v) | 50 mL |
| Leucine 1% | 0.01% | 10 mL |
| Adenine 0.2% | 0.002% | 10 mL |
| Histidine 1% | 0.002% | 2 mL |
| Tryptophan 1% | 0.002% | 2 mL |
| Methionine 1% | 0.002% | 2 mL |
| Lysine 1% | 0.003% | 3 mL |
| Water | n/a | Up to 1,000 mL |
Sterilize by autoclaving. The amino acid composition may vary depending on the yeast strain used. Store at room temperature (RT).
2. Synthetic minimal glycerol (SMGly)
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| Yeast nitrogen base | 1.7 g/L | 1.7 g |
| Ammonium sulfate | 5 g/L | 5 g |
| Glycerol 30% (v/v) | 3% (v/v) | 100 mL |
| Leucine 1% | 0.01% | 10 mL |
| Adenine 0.2% | 0.002% | 10 mL |
| Histidine 1% | 0.002% | 2 mL |
| Tryptophan 1% | 0.002% | 2 mL |
| Methionine 1% | 0.002% | 2 mL |
| Lysine 1% | 0.003% | 3 mL |
| Water | n/a | Up to 1,000 mL |
Sterilize by autoclaving. The amino acid composition may vary depending on the yeast strain used. Store at RT.
3. Buffer SHP
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| Sorbitol 1.2 M | 0.6 M | 5 mL |
| HEPES 100 mM | 20 mM | 2 mL |
| PMSF 200 mM | 1 mM | 50 μL |
| Complete Mini, EDTA-free | 1 tablet | 1 tablet |
| Total | n/a | 10 mL |
Prepare PMSF fresh with absolute ethanol. Complete Mini, EDTA-free is stored at 4 °C. Buffer SHP should be prepared freshly before use.
4. TBS 10×
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| Trizma base | 500 mM | 121.14 g |
| NaCl | 1.5 M | 175.32 g |
| Adjust pH to 7.6 with HCl | … | … |
| Total | n/a | 2,000 mL |
Measure pH using a calibrated pH meter. Store at RT.
5. TBS-Tween 10×
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| Trizma base | 500 mM | 121.14 g |
| NaCl | 1.5 M | 175.32 g |
| Tween | 0.1% (v/v) | 2 mL |
| Adjust pH to 7.6 with HCl | … | … |
| Total | n/a | 2,000 mL |
Measure pH using a calibrated pH meter. Store at RT.
6. LAE 2×
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| SDS | 4% | 0.4 g |
| Glycerol | 20% (v/v) | 2 mL |
| β-mercaptoethanol | 10% (v/v) | 1 mL |
| Bromophenol blue | 0.04% | 4 mg |
| Tris-HCl pH 6.8 1 M | 0.120 M | 1.2 mL |
| Total | n/a | 10 mL |
β-mercaptoethanol is stored at 4 °C. Measure pH using a calibrated pH meter. Store LAE 2× at -20 °C.
7. SDS-PAGE: running gel 14% (1 mm thickness)
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| Acrylamide 30%, Bis-acrylamide 0.2% | 14% (w/v) | 2.55 mL |
| Tris 8.75 M, pH 8.8 | 375 mM | 1.17 mL |
| Distilled water (H2O) | n/a | 1.69 mL |
| SDS 10% | 0.1% (w/v) | 55 μL |
| APS 10% | 0.05% (w/v) | 33 μL |
| TEMED | 0.05% (v/v) | 3.30 μL |
| Total | n/a | 5.5 mL |
Acrylamide 30%, Bis-acrylamide 0.2% is stored at 4 °C. Measure pH of Tris using a calibrated pH meter.
8. SDS-PAGE: stacking gel 4.85% (1 mm thickness)
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| Acrylamide 30%, Bis-acrylamide 0.2% | 4.85% (w/v) | 0.32 mL |
| Tris 0.6 M, pH 6.8 | 60 mM | 0.2 mL |
| Distilled water (H2O) | n/a | 1.45 mL |
| SDS 10% | 0.1% (w/v) | 20 μL |
| APS 10% | 0.05% (w/v) | 10 μL |
| TEMED | 0.1% (v/v) | 2 μL |
| Total | n/a | 2.0 mL |
Acrylamide 30%, Bis-acrylamide 0.2% is stored at 4 °C. Measure pH of Tris using a calibrated pH meter.
9. Running buffer 10×
| Reagent | Final concentration | Quantity or volume |
|---|---|---|
| Trizma base | 0.25 M | 60.8 g |
| Glycine | 1.92 M | 288 g |
| SDS | 1% (w/v) | 20 g |
| Total | n/a | 2,000 mL |
Store at RT.
Laboratory supplies
1. Glass beads (Sigma, catalog number: G9268)
2. Microtubes 2 mL (Axygen, catalog number: MCT-200-C)
3. Microtubes 1.5 mL (CRM Globe, catalog number: PD1010)
4. Cuvettes (Sarstedt, catalog number: 67.742)
5. Nitrocellulose membrane 0.2 μm (Amersham Protran, Cytiva, catalog number: 10600015)
6. 10 μL pipette tips (Sarstedt, catalog number: 70.3030.020)
7. 200 μL pipette tips (Sarstedt, catalog number: 70.3030.020)
8. 1,000 μL pipette tips (Sarstedt, catalog number: 70.3050.020)
9. Culture flasks 250 mL (Duran, catalog number: 090961)
10. Conical tubes 50 mL (Sarstedt, catalog number: 62.547.254)
Equipment
1. Refrigerated centrifuge (Haier, model: LX-165T2R)
2. Refrigerated centrifuge (Beckman Coulter, model: Alegra AL2086003)
3. Rotary incubator shaker (New Brunswick SC, model: INNOVA44)
4. Vortex (Scientific industries, Inc., model: G560)
5. Spectrophotometer (Eppendorf BioPhotometer Plus, model: 6132)
6. Electrophoresis chamber (Bio-Rad, model: Mini PROTEAN Tetra Cell, catalog number: 552BR019843)
7. Semi-dry chamber (Bio-Rad, model: Trans-Blot SD Cell, catalog number: 221BR56787)
8. pH meter (Nanbei, model: PH200)
Procedure
文章信息
稿件历史记录
提交日期: Dec 5, 2025
接收日期: Jan 18, 2026
在线发布日期: Jan 28, 2026
出版日期: Feb 20, 2026
版权信息
© 2026 The Author(s); This is an open access article under the CC BY-NC license (https://creativecommons.org/licenses/by-nc/4.0/).
如何引用
Mendoza-Martínez, A. E., Bravo-Arévalo, J. E., Pedroza-Dávila, U. and Funes, S. (2026). Optimized Mechanical Isolation of Mitochondria From Saccharomyces cerevisiae Preserving Atg32 for Quantitative Analysis. Bio-protocol 16(4): e5610. DOI: 10.21769/BioProtoc.5610.
分类
细胞生物学 > 细胞器分离 > 线粒体
细胞生物学 > 细胞信号传导 > 自体吞噬
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。