

Deaminase-Assisted Sequencing for the Identification of 5-glyceryl-methylcytosine
脱氨酶辅助测序用于鉴定 5-甘油基甲基胞嘧啶
发布: 2026年02月20日第16卷第4期 DOI: 10.21769/BioProtoc.5601 浏览次数: 403
评审: Anonymous reviewer(s)
参见作者原研究论文
The authors used this protocol in:

Oct 2025
Advertisement











Abstract
DNA epigenetic modifications play crucial roles in regulating gene expression and cellular function across diverse organisms. Among them, 5-glyceryl-methylcytosine (5gmC), a unique DNA modification first discovered in Chlamydomonas reinhardtii, represents a novel link between redox metabolism and epigenetic regulation. Accurate genome-wide detection of 5gmC is essential for investigating its biological functions, yet no streamlined method has been available. Here, we present deaminase-assisted sequencing (DEA-seq), a simple and robust approach for base-resolution mapping of 5gmC. DEA-seq employs a single DNA deaminase that efficiently converts unmodified cytosines (C) and 5-methylcytosine (5mC) into uracils or thymines, while leaving 5gmC intact. This selective resistance generates a clear sequence signature that enables precise identification of 5gmC sites across the genome. The method operates under mild reaction conditions and is compatible with low-input DNA, minimizing sample loss and improving detection sensitivity. Overall, DEA-seq provides an accessible, efficient, and highly accurate protocol for profiling 5gmC, offering clear advantages in workflow simplicity, DNA integrity, and analytical performance.
Key features
• A commercial deaminase mix (DEA) efficiently converts unmodified cytosines and 5-methylcytosines into uracils or thymines.
• 5gmC specifically resists DEA-mediated deamination, enabling its identification at single-base resolution.
• DEA-seq requires only minimal DNA input and supports high-sensitivity detection from nanogram-level samples.
Keywords: 5-glyceryl-methylcytosine (5gmC) (5-甘油基甲基胞嘧啶)Graphical overview
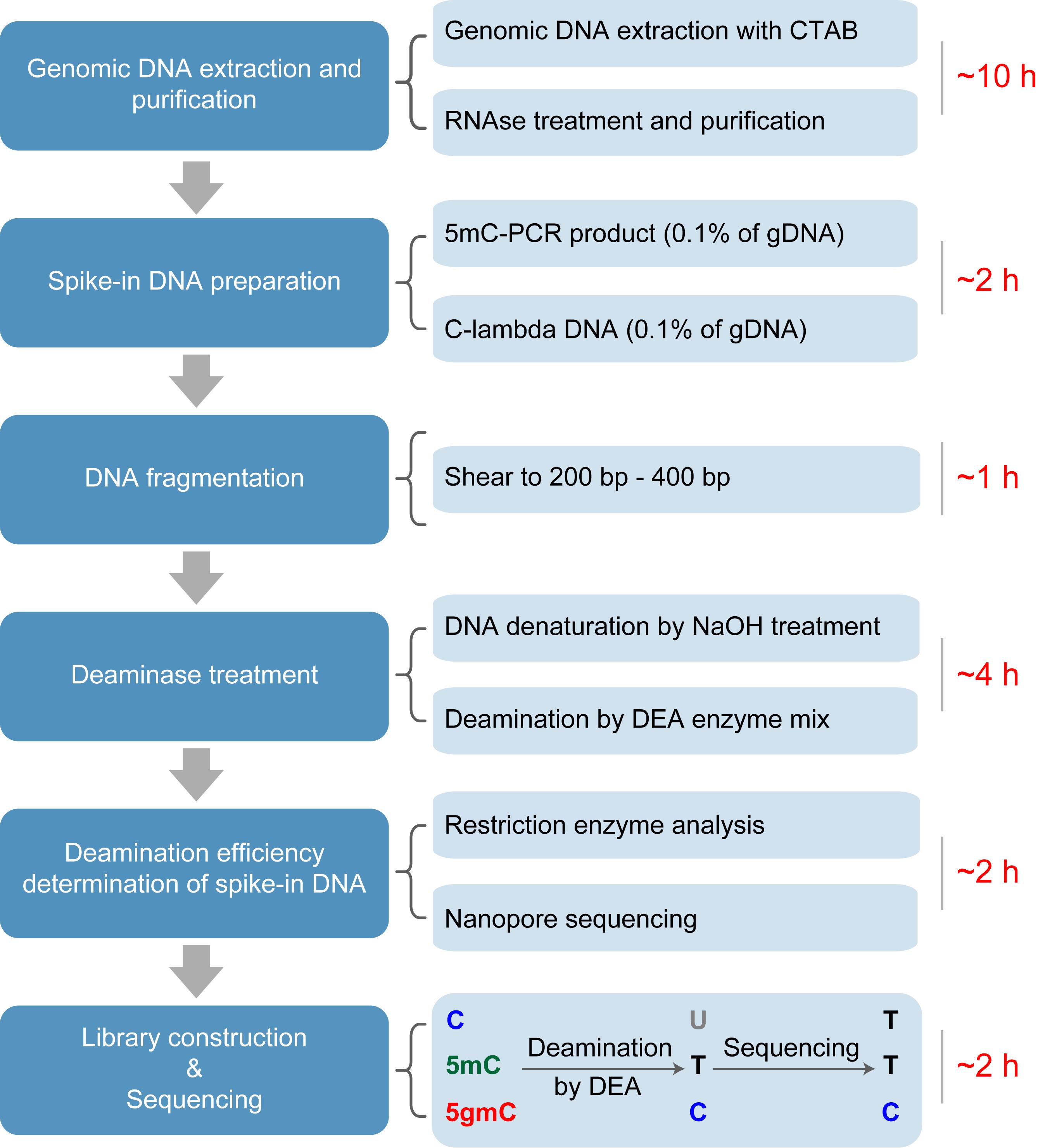
The workflow of deaminase-assisted sequencing (DEA-seq). Genomic DNA is extracted from Chlamydomonas reinhardtii following a standard protocol. After RNA removal using an RNase A/T1 mix, the DNA is further purified by ethanol precipitation. Unmodified lambda DNA and PCR-amplified 5mC-DNA are then added as spike-ins, followed by ultrasonic shearing of the mixture into ~300 bp fragments. The fragmented DNA is denatured into single-stranded DNA (ssDNA) using NaOH treatment. After deaminase treatment, unmodified cytosines and 5mC are converted into uracils and thymines, respectively, while 5gmC remains resistant to deamination. Following PCR amplification and sequencing, both U and T are read as T, whereas 5gmC is retained as C, enabling precise identification of 5gmC at single-base resolution.
Background
DNA methylation in eukaryotes primarily occurs at the fifth carbon of cytosines (C), forming 5-methylcytosine (5mC). In mammals, ten-eleven translocation (TET) enzymes catalyze the stepwise oxidation of 5mC to produce 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC), and 5-carboxylcytosine (5caC), which can contribute to both active and passive DNA demethylation [1]. In the unicellular green alga Chlamydomonas reinhardtii, a TET homologue named 5mC modifying enzyme 1 (CMD1) catalyzes a distinct reaction that converts 5mC into 5-glyceryl-methylcytosine (5gmC) using vitamin C, rather than α-ketoglutarate, as a co-substrate [2,3]. This unusual enzymatic activity not only expands the repertoire of cytosine modifications in eukaryotic genomes but also provides a direct mechanistic link between redox metabolism and epigenetic regulation. 5gmC may function as an intermediate in DNA demethylation and plays an essential role in the feedback regulation of photosynthesis in C. reinhardtii [2]. Importantly, mass spectrometry–based analyses have shown that 5mC and 5gmC are the only cytosine modifications detected in the C. reinhardtii genome, with no evidence for the presence of oxidized cytosine derivatives such as 5hmC, 5fC, or 5caC [2,4]. However, its genome-wide distribution and potential function as an independent epigenetic mark remain poorly understood. Accurate detection of 5gmC is crucial for functional studies but remains technically challenging because this modification does not alter Watson–Crick base pairing and therefore cannot be distinguished from unmodified cytosines or 5mC using conventional sequencing approaches.
Bisulfite sequencing (BS-seq) is widely considered the gold standard for base-resolution mapping of 5mC, but its harsh chemical treatment causes substantial DNA degradation and compromises sequencing quality [5]. More importantly, BS-seq cannot discriminate among different cytosine modifications, such as 5mC and 5hmC. Several BS-seq-derived methods have been developed to address this limitation. For example, TET-assisted bisulfite sequencing (TAB-seq) profiles 5hmC by protecting it with β-glucosyltransferase (β-GT) and selectively deaminating 5mC and unmodified C through TET oxidation followed by bisulfite treatment [6]. Other methods independent of BS-seq have also been established, including TET-assisted pyridine borane sequencing (TAPS) for detecting 5mC and 5hmC [7] and APOBEC-coupled epigenetic sequencing (ACE-seq), which enables reliable identification of 5hmC [8,9]. However, these techniques typically involve multiple enzymatic or chemical steps and may suffer from incomplete conversion or false-positive signals, particularly when detecting rare DNA modifications. Previously, a TET-coupled bisulfite sequencing (TET-BS-seq) approach was developed to distinguish 5gmC from 5mC, taking advantage of the resistance of 5gmC to both TET-mediated oxidation and bisulfite deamination [2]. Nevertheless, given the extremely low abundance of 5gmC in the genome (~10 ppm of total cytosines) [10], the accuracy and sensitivity of TET-BS-seq remain limited.
To address these challenges, we developed deaminase-assisted sequencing (DEA-seq), a simplified and efficient method for genome-wide identification of 5gmC [4]. This approach employs a commercial DNA deaminase enzyme mix (DEA), which is capable of converting unmodified C and 5mC into uracils or thymines, while leaving 5gmC unaffected. The selective resistance of 5gmC to DEA-mediated deamination mirrors the chemical protection observed for 5-glycosyloxymethylcytosine (5ghmC) in ACE-seq [8], enabling precise discrimination of 5gmC in a single-step reaction. DEA-seq thus provides a robust and streamlined protocol for detecting 5gmC at base resolution, facilitating further exploration of its biological functions and regulatory mechanisms in C. reinhardtii.
Materials and reagents
Biological materials
1. Chlamydomonas reinhardtii strains (CC-125, wild type mt+; and CC-5325, cw15 mt-)
Reagents
1. TaKaRa TaqTM Hot Start Version (Takara, catalog number: R007A)
2. RNase A/T1 mix (Thermo Fisher Scientific, catalog number: EN0551)
3. HpyCH4IV (TaiI) (NEB, catalog number: R0619S)
4. FastDigest SsiI (AciI) (Thermo Fisher Scientific, catalog number: FD1794)
5. 5-methyl-dCTP (2.5 mM) (NEB, catalog number: N0356S)
6. dATP (100 mM) (Takara, catalog number: 4026)
7. dTTP (100 mM) (Takara, catalog number: 4029)
8. dGTP (100 mM) (Takara, catalog number: 4027)
9. NaOH (Macklin, catalog number: S817971)
10. Agarose (BBI, catalog number: A620014)
11. 100 bp DNA ladder (TransGen Biotech, catalog number: BM301-01)
12. 1 kb DNA ladder (TransGen Biotech, catalog number: BM201-01)
13. 10× loading buffer (Takara, catalog number: 9157)
14. lambda DNA (dam-, dcm-) (Takara, catalog number: 3019)
15. DNA Clean & Concentrator-5 (Zymo Research, catalog number: D4013) (contains DNA binding buffer, DNA wash buffer, DNA elution buffer, Zymo-SpinTM columns, and collection tubes)
16. EpiArt DNA Enzymatic Methylation kit (Vazyme, catalog number: EM301) (contains oxidation reaction buffer, oxidation reaction S, oxidation reagent, oxidation enzyme 1, oxidation enzyme 2, active reagent, stop reagent, DEA reaction buffer, BSA, DEA enzyme mix, and dilution buffer)
17. EpiArt DNA Methylation Library kit for Illumina V3 (Vazyme, catalog number: NE103) (contains 3′ ligation buffer, 3′ ligation enzyme mix, 3′ adapter, extension primer, extension enzyme mix, 5′ ligation mix, 5′ adapter, and VAHTS HiFi Amplification Mix V3)
18. VAHTS multiplex oligos set 4/5 for Illumina (Vazyme, catalog number: N321/N322-01)
19. QubitTM ssDNA Assay kit (Thermo Fisher Scientific, catalog number: Q10212) (contains QubitTM ssDNA reagent, QubitTM ssDNA buffer, QubitTM ssDNA standard #1, and QubitTM ssDNA standard #2)
20. SanPrep Column DNA Gel Extraction kit (Sangon, catalog number: B518131) (contains buffer B2, wash solution, elution buffer, spin columns, and collection tubes)
21. Tris (Solarbio, catalog number: T8060)
22. Glacial acetic acid (Solarbio, catalog number: A801295)
23. EDTA (Solarbio, catalog number: E8040)
24. Phenol-chloroform (Meryer, catalog number: N09169-500ML)
25. Ethanol (Sinopharm, catalog number: 10009259)
26. Chloroform (Sinopharm, catalog number: 10006818)
27. Isopropanol (Sinopharm, catalog number: 80109218)
Note: All reagents should be stored according to the manufacturer’s instructions. The DEA enzyme mix is critical for this protocol because it exhibits specific deamination activity toward unmodified cytosine and 5mC, but not 5gmC. In contrast, other reagents may be substituted with suitable alternatives, provided that equivalent performance is validated.
Solutions
1. 50× TAE and 1× TAE (see Recipes)
Recipes
1. 50× TAE and 1× TAE
a. Dissolve 242 g of Tris base in deionized water and add 57.1 mL of glacial acetic acid and 100 mL of 0.5 M EDTA (pH 8.0). Adjust the final volume to 1 L with deionized water and mix thoroughly.
b. Dilute the 50× TAE stock solution with deionized water to prepare 1× TAE working solution.
Laboratory supplies
1. QubitTM assay tubes (Thermo Fisher Scientific, catalog number: Q32856)
2. 1.5 mL tubes (QINI, catalog number: CN.0100015S)
3. 0.2 mL PCR tubes (QINI, catalog number: CN.0100002S)
Equipment
1. Pipettes (2.5 μL, 20 μL, 200 μL and 1,000 μL) (Eppendorf)
2. M220 Focused ultrasonicator (Covaris, catalog number: M220)
3. NanoDrop One (Thermo Fisher Scientific, catalog number: 840-317400)
4. ProFlexTM 3 × 32-well PCR system (Thermo Fisher Scientific, catalog number: 4484073)
5. QubitTM 4 fluorometer (Thermo Fisher Scientific, catalog number: Q33238)
6. VAHTS DNA clean beads (Vazyme, catalog number: N411-01)
7. Centrifuge (Eppendorf, model: Centrifuge 5425/5425 R)
8. Mini centrifuge (Joanlab, model: MC-4K)
9. Vortex mixer (Joanlab, model: VM-400-VM-400Max)
10. Gel image analysis system (Tanon, catalog number: 21TMINI1000-11141)
11. Electrophoresis system (Tanon, catalog number: EPS600)
12. Scalpels (Leica, catalog number: DB80LX)
Software and datasets
1. Trimmomatic (v0.39, http://www.usadellab.org/cms/?page=trimmomatic)
2. Bismark (v0.24.2, https://felixkrueger.github.io/Bismark/)
Procedure
文章信息
稿件历史记录
提交日期: Nov 24, 2025
接收日期: Jan 13, 2026
在线发布日期: Jan 20, 2026
出版日期: Feb 20, 2026
版权信息
© 2026 The Author(s); This is an open access article under the CC BY-NC license (https://creativecommons.org/licenses/by-nc/4.0/).
如何引用
He, B. D., Wang, F. C. and Xue, J. H. (2026). Deaminase-Assisted Sequencing for the Identification of 5-glyceryl-methylcytosine. Bio-protocol 16(4): e5601. DOI: 10.21769/BioProtoc.5601.
分类
生物信息学与计算生物学
分子生物学 > DNA > DNA 修饰
分子生物学 > DNA > DNA 测序
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。