

A New Approach to Generate Gastruloids to Develop Anterior Neural Tissues
一种产生原肠胚样细胞以发育前神经组织的新方法
发布: 2023年07月20日第13卷第14期 DOI: 10.21769/BioProtoc.4722 浏览次数: 3295
评审: Rajesh RanjanHélène LégerSamantha Haller
参见作者原研究论文
The authors used this protocol in:

May 2021
Advertisement








Abstract
Embryonic development is a complex process integrating cell fate decisions and morphogenesis in a spatiotemporally controlled manner. Previous studies with model organisms laid the foundation of our knowledge on post-implantation development; however, studying mammalian embryos at this stage is a difficult and laborious process. Early attempts to recapitulate mammalian development in vitro begun with embryoid bodies (EBs), in which aggregates of mouse embryonic stem cells (mESCs) were shown to differentiate into spatially arranged germ layers. A more revised version of EBs, gastruloids, improved the germ layer differentiation efficiency and demonstrated cell fate patterning on multiple axes. However, gastruloids lack anterior neural progenitors that give rise to brain tissues in the embryo. Here, we report a novel culture protocol to coax mESCs into post-implantation epiblast-like (EPI) aggregates in high throughput on bioengineered microwell arrays. We show that upon inhibition of the Wnt signaling pathway, EPI aggregates establish an extended axial patterning, leading to co-derivation of anterior neural progenitors and posterior tissues. Our approach is amenable to large-scale studies aimed at identifying novel regulators of gastrulation and anterior neural development that is currently out of reach with existing embryoid models. This work should contribute to the advancement of the nascent field of synthetic embryology, opening up exciting perspectives for various applications of pluripotent stem cells in disease modeling and tissue engineering.
Key features
• A new gastruloid culture system to model post-implantation mouse embryonic development in vitro
• High-throughput formation of epiblast-like aggregates on hydrogel microwells
• Builds upon conventional gastruloid cultures and provides insight into the role of Wnt signaling for the formation of anterior neural tissues
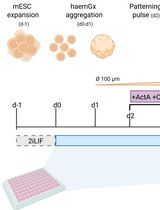
Graphical overview

Background
The first attempts to model mouse embryonic development in vitro begun with a coincidental observation. When teratocarcinomas from neonatal mouse testes were analyzed, scientists noticed a structural similarity to the developing mouse embryo (Stevens, 1959). Hence, they called these tumors “embryoid bodies” (Pierce et al., 1960). It was further shown that embryoid bodies retained the tumor-forming capacity to form cell types ranging from cartilage to neural tissue, a multipotency that was later attributed to the embryonal carcinoma (EC) cells (Kleinsmith and Pierce, 1964). The use of EC cells to mimic embryonic development was rapidly replaced by embryonic stem cells (ESCs) following their isolation in 1981 (Evans and Kaufman, 1981). Pioneering studies have demonstrated self-organization potential of embryoid bodies to recapitulate, to a limited extent, gastrulation-like events and antero-posterior axis determination (ten Berge et al., 2008). However, this was not accompanied by axial morphogenesis.
More recently, gastruloids have taken the extent of self-organization potential of mouse ESCs (mESCs) to demonstrate that embryoid bodies could undergo axial morphogenesis (van den Brink et al., 2014). In this model, when treated with Wnt agonist CHIR99021, small aggregates of mESCs (~300 cells) were shown to break symmetry and demonstrated polarized T/Bra expression in a reproducible way. When cultured further, gastruloids elongated and established patterning along antero-posterior, dorso-ventral, and medio-lateral axes (Beccari et al., 2018). Moreover, the multi-axial patterning was linked to spatiotemporal activation of Hox gene clusters, a phenomenon that is conserved across many species (Santini et al., 2003). Such self-organization potential of mESCs, in the absence of any extraembryonic tissue, was remarkably similar to the developing post-occipital region of the mouse embryo; however, tissues mapping to anterior brain regions were completely absent in gastruloids.
Mechanical forces and extracellular matrix composition have significant influence on the developing mouse embryo (Hiramatsu et al., 2013). When gastruloids are embedded in a basement membrane substitute, they remarkably organize into structures bearing somites and a central neural tube-like tissue (Veenvliet et al., 2020). Even under these conditions, gastruloids fail to form any anterior neural tissues corresponding to the brain regions.
The epiblast is the domain that forms the embryo proper. However, contribution from extraembryonic tissues is required since development is halted in their absence (Donnison et al., 2005; Rodriguez et al., 2005). Studies accounting for this necessity have therefore established hybrid embryoid models, combining embryonic stem cells with trophoblast stem cells (TSCs) (Harrison et al., 2017; Rivron et al., 2018) and/or extraembryonic endoderm cells (XENs) (Sozen et al., 2018). More recently, mESCs were co-cultured with transdifferentiated TSCs and XENs to generate structures that almost completely recapitulate E8.5 embryos, including brain tissues (Amadei et al., 2022; Tarazi et al., 2022). However, the complexity of the tri-culture system and necessity of special equipment to grow them until late stages remain as the major limitations in generating synthetic embryos.
Previously, it was shown that embryos lacking extraembryonic Wnt source could still break symmetry and initiate gastrulation, suggesting an autonomous developmental potential of epiblast cells (Yoon et al., 2015). Moreover, overactivation of Wnt signaling in the epiblast was shown to deplete anterior neural progenitors in the favor of mesoderm derivatives; this phenotype could be rescued by inhibition of Wnt signaling (Osteil et al., 2019). Therefore, we reasoned that a similar trade-off could be happening in conventional gastruloids, owing to overactivation of Wnt signaling by CHIR99021 treatment.
Here, we report a new model system based on aggregation of mESCs to derive post-implantation epiblast-like structures (EPI aggregates). We formulated a serum-free epiblast-induction medium comprising Activin-A (Tgf-β agonist), Fgf2 (Fgf agonist), and knockout serum replacement, which promoted the acquisition of epiblast identity, followed by their spontaneous symmetry breaking and subsequent morphogenesis without any external Wnt stimulation. Moreover, inhibition of Wnt signaling during early stages of EPI aggregate formation helped to maintain anterior neural progenitors, which then committed to generate forebrain-, midbrain-, and hindbrain-like tissues. This protocol expands the cell type repertoire that can be generated within gastruloids.
Materials and reagents
Mouse embryonic stem cell lines
SBr [SOX1-GFP; T/BRA-mCherry] mESC line (Deluz et al., 2016)
WNT [TLC-mCherry] mESC line (Ferrer-Vaquer et al., 2010; Faunes et al., 2013)
TGF-β [AR8-mCherry] mESC line (Serup et al., 2012)
Cell culture materials
Ultra-low attachment 96-well plate (Corning, catalog number: 7007)
6-well plate, tissue culture–treated (Falcon, catalog number: 38016)
6-well plate, non tissue culture–treated (Corning, catalog number: CLS3471)
PEG microwells (400 μm microwell diameter, 121 wells/array in 24-well format) (Brandenberg et al., 2020). Commercially available Gri3D® 96-well plate (400 μm microwell diameter, 121 wells/array) could be used alternatively.
10 cm plate, non tissue culture–treated (Corning, catalog number: CLS430591)
Pipettes (2, 5, 10, 25, and 50 mL)
Micropipettes (1–10, 2–20, 20–200, and 100–1,000 μL)
Pipette tips (10, 20, 200, and 1,000 μL)
Cell culture reagents
Base media and dissociation reagents
DMEM + GlutaMAX (Gibco, catalog number: 61965-026). Keep at 4 °C
DMEM/F12 + GlutaMAX (Gibco, catalog number: 31331-028). Keep at 4 °C
Neurobasal (Gibco, catalog number: 21103-049). Keep at 4 °C
Accutase (Gibco, catalog number: A11105). Keep at 4 °C
PBS (Gibco, catalog number: 10010023). Keep at room temperature
Media supplements
ESC-qualified FBS (Gibco, catalog number: 16141-079). Keep stock at -80 °C, working aliquots at -20 °C
KnockOutTM serum replacement (Thermo Fisher Scientific, catalog number: 10828010). Keep stock at -80 °C, working aliquots at -20 °C
N2 supplement (Gibco, catalog number: 17502001). Keep at -20 °C
B27 supplement (Gibco, catalog number: 17504001). Keep at -20 °C
Sodium pyruvate (Gibco, catalog number: 11360-070). Keep at 4 °C
MEM non-essential amino acids (Gibco, catalog number: 11140-035). Keep at 4 °C
2-mercaptoethanol (Gibco, catalog number: 31350-010). Keep at 4 °C
Penicillin/Streptomycin (Gibco, catalog number: 15140-122). Keep working aliquots at -20 °C
GlutaMAX (Gibco, catalog number: 35050-038). Keep at room temperature
Growth factors and small molecule inhibitors
CHIR99021 (Tocris, catalog number: 4423). Keep stock at -80 °C, working aliquots at 4 °C
PD0325901 (Selleckchem, catalog number: S1036). Keep stock at -80 °C, working aliquots at 4 °C
LIF (in-house preparation). Keep stock at -80 °C, working aliquots at 4 °C
FGF2 (Gibco, catalog number: PMG0035). Keep stock at -80 °C, working aliquots at -20 °C. Use within three months after reconstitution
ACTIVIN A (R&D systems, catalog number: 338-AC). Keep stock at -80 °C, working aliquots at -20 °C. Use within three months after reconstitution
XAV939 (Tocris, catalog number: 3748). Keep stock at -80 °C, working aliquots at -20 °C
Immunostaining materials and reagents
4% PFA (Thermo Fisher, catalog number: J19943.K2)
DAPI (Sigma, catalog number: 9542)
Glass slides
Coverslips (1.5 thickness)
Mounting medium (Vectashield, catalog number: H-1000-10)
Nail polish
Triton X-100 (Sigma, catalog number: X100)
Mouse embryonic stem cell (mESC) maintenance medium (500 mL) (see Recipes)
EPI differentiation medium (500 mL) (see Recipes)
Primary and secondary antibodies (Table 1)
Table 1. List of primary and secondary antibodies
| Target | Species | Dilution | Catalogue number | Supplier |
|---|---|---|---|---|
| Anti-E-cadherin | Rabbit | 1:500 | 24E10 | Cell Signaling Technology |
| Anti-Sox2 | Rabbit | 1:400 | ab97959 | Abcam |
| Anti-Sox1 | Goat | 1:50 | af3369 | R&D Systems |
| Anti-Otx2 | Goat | 1:25 | af1979 | R&D Systems |
| Anti-Brachyury | Goat | 1:300 | sc-17745 (C-19) | Santa Cruz |
| Anti-Brachyury | Rabbit | 1:100 | ab209665 | Abcam |
| Anti-Oct4 | Mouse | 1:200 | sc-5270 (C-10) | Santa Cruz |
| Anti-Nanog | Rat | 1:300 | 14-5761-80 | Thermo Fisher |
| Anti-Cdx2 | Rabbit | 1:200 | ab76541 | Abcam |
| Anti- aPKC | Mouse | 1:100 | sc-17781 (H-1) | Santa Cruz |
| Anti-Sox17 | Goat | 1:200 | AF1924 | Abcam |
| Anti-Cdx2 | Rabbit | 1:200 | ab76541 | Abcam |
| Anti-mCherry | Rat | 1:400 | M11217 | Thermo Fisher |
| Phalloidin AF488 | 1:1,000 | A12379 | Thermo Fisher | |
| Phalloidin AF635 | 1:1,000 | A34054 | Thermo Fisher | |
| Anti-chicken Alexa Fluor 488 | Donkey | 1:500 | 703-545-155 | Jackson ImmunoResearch |
| Anti-rat Alexa Fluor 568 | Goat | 1:500 | A-21247 | Thermo Fisher |
| Anti-goat Alexa Fluor 568 | Donkey | 1:500 | A-11057 | Thermo Fisher |
| Anti-goat Alexa Fluor 647 | Donkey | 1:500 | A-21447 | Thermo Fisher |
| Anti-rabbit Alexa Fluor 488 | Donkey | 1:500 | A-21206 | Thermo Fisher |
| Anti-rabbit Alexa Fluor 568 | Donkey | 1:500 | A-10042 | Thermo Fisher |
| Anti-rabbit Alexa Fluor 647 | Donkey | 1:500 | A-31573 | Thermo Fisher |
| Anti-mouse Alexa Fluor 568 | Donkey | 1:50 | A-10037 | Thermo Fisher |
| Anti-mouse Alexa Fluor 647 | Donkey | 1:50 | A-31571 | Thermo Fisher |
Equipment
Incubator with regulated temperature and humidity
Centrifuge
Biological safety cabinet
4 °C fridge, -20 °C freezer, and -80 °C freezer
Eclipse inverted microscope (Nikon, catalog number: TS100)
Pipettes (single and multichannel)
Sterile tips
Hemocytometer and Countess 3 automated cell counter (catalog number: AMQAX2000)
Zeiss LSM700 inverted confocal microscope
Nikon Eclipse Ti inverted microscope
EC Plan-Neofluar 10×/0.30 and Plan-Apochromat 20×/0.80 air objectives
Software
Zen software (2009)
NIS elements (version 4.11.0)
Fiji ImageJ (version 2.0.0-rc-69/1.52n)
GraphPad Prism software (version 8.4.2)
RStudio (version 1.3.1056)
Procedure
文章信息
版权信息
© 2023 The Author(s); This is an open access article under the CC BY-NC license (https://creativecommons.org/licenses/by-nc/4.0/).
如何引用
Girgin, M., Broguiere, N., Mattolini, L. and Lutolf, M. (2023). A New Approach to Generate Gastruloids to Develop Anterior Neural Tissues. Bio-protocol 13(14): e4722. DOI: 10.21769/BioProtoc.4722.
分类
干细胞 > 胚胎干细胞 > 细胞分化
发育生物学 > 形态建成 > 器官形成
细胞生物学 > 细胞成像 > 活细胞成像
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。