

Purification of Mitochondrial Ribosomes with the Translocase Oxa1L from HEK Cells
用转位酶Oxa1L纯化HEK细胞线粒体核糖体
(*contributed equally to this work) 发布: 2021年08月05日第11卷第15期 DOI: 10.21769/BioProtoc.4110 浏览次数: 5293
评审: Khyati Hitesh ShahAndrea SaponaroAnonymous reviewer(s)

参见作者原研究论文
The authors used this protocol in:

Nov 2020
Advertisement









Abstract
Mitochondrial ribosomes (mitoribosomes) perform protein synthesis inside mitochondria, the organelles responsible for energy conversion and adenosine triphosphate (ATP) production in eukaryotic cells. To investigate their functions and structures, large-scale purification of intact mitoribosomes from mitochondria-rich animal tissues or HEK cells have been developed. However, the fast purification of mitoribosomes anchored to the mitochondrial inner membrane in complex with the Oxa1L translocase remains particularly challenging. Herein, we present a protocol recently developed and modified in our lab that provides details for the efficient isolation of intact mitoribosomes with its translocase Oxa1L. We combined the cell culture of PDE12-/- or wild-type HEK293 cell lines with the isolation of mitochondria and the purification steps used for the biochemical and structural studies of mitoribosomes and Oxa1L.
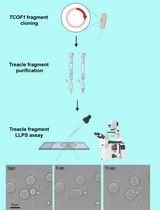
Graphic abstract:

Schematic procedure for the purification of mitoribosomes from HEK cells. The protocol described herein includes two main sections: 1) isolation of mitochondria from HEK cells; and 2) purification of mitoribosome-Oxa1L from mitochondria. RB: Resuspension Buffer (see Recipes) (Created with BioRender.com).
Background
Human mitochondria possess their own genome and house all the components necessary for transcription, RNA maturation, and translation of the encoded genetic information (Dennerlein et al., 2017). The mitoribosome, one of the most important machineries within mitochondria, is responsible for the translation of essential mitochondrial mRNAs encoding components of oxidative phosphorylation (OXPHOS) complexes (Ott et al., 2016). Since all proteins translated by the human mitoribosome are membrane proteins, knowledge of the mechanism of co-translational membrane protein insertion is central to understanding mitoribosome translation (Hildenbeutel et al., 2012; Stiller et al., 2016). In human mitochondria, the oxidase assembly 1-like (Oxa1L) translocase plays a central role in the membrane insertion of mitochondrially encoded products (Haque et al., 2010; Stiller et al., 2016); however, due to the unique features of mitochondria, little is known regarding the molecular and quality control mechanisms underlying mitochondrial translation.
Owing to the structures obtained by single-particle electron cryo-microscopy (cryo-EM), opportunities now arise to comprehensively study the molecular mechanisms underlying the translational action and quality control of the human mitoribosome (Rathore, 2020). Although the existence of quality control in mitochondria has been predicted based on analogy with bacterial and eukaryotic cytosols (Ayyub et al., 2020), biochemical or structural evidence of ribosome-associated quality control in mitochondria remain elusive. We reasoned that any attempt to induce translational stalling may trap mitoribosomes in various stages of the translation cycle during the act of nascent chain insertion into the inner mitochondrial membrane, and generate intermediates suitable for structural analysis by cryo-EM. To this end, we purified mitoribosomes from a genetically engineered human cell line lacking the 2’-5’ phosphodiesterase exonuclease 12 (PDE12). PDE12 facilitates the maturation of mt-tRNA; therefore, a PDE12 knockout can lead to aberrant polyadenylation of the 3’ ends of mt-tRNAs and, consequently, mitoribosome stalling (Rorbach et al., 2011; Pearce et al., 2017). As expected, we observed a substantial proportion of stalled mitoribosomes and discovered a mitoribosome-associated quality control pathway (mtRQC) to rescue the elongational stalling ribosomes (Desai et al., 2020). Our report of the first mitoribosome-associated quality control pathway provides novel insights into the regulation of mitochondrial translation. Since the cell needs to detect and respond rapidly even to subtle changes in translation rates, quality control is intimately coupled to elongation. During purification, we included GMPPCP, a non-hydrolysable GTP analog, to prevent dissociation of GTPases from mitoribosomes, and successfully captured five additional structures of virtually every elongating ribosomal state. Purifying the mitoribosome in complex with its translocase Oxa1L was challenging since membrane proteins are unstable and tend to dissociate during complicated isolation steps. Accordingly, we added n-dodecyl β-D-maltoside (β-DDM) along with cardiolipin to solubilize mitoribosomes anchored to the mitochondrial inner membrane via the Oxa1L translocase. We also trapped Oxa1L on all our active and stalled ribosomes, giving us a first glimpse at the co-translational insertion of mitochondrial inner membrane proteins.
This protocol was modified from prior methods developed in our lab (Amunts et al., 2015; Brown et al., 2017) and by others (Greber et al., 2014 and 2015; Aibara et al., 2018), as well as in our current work (Desai et al., 2020). The method described here can be used to obtain mitoribosomes in complex with Oxa1L, which has advantages such as the applicability for mitochondria isolation from large volumes of suspension cell culture, the use of common media and buffers, ease of handling, and absence of additional affinity purification steps. In our previous publication and experiments, we observed a good reproducibility between HEK293 cell batches using this methodology.
Materials and Reagents
T75/T175 flasks (Thermo ScientificTM)
1 L/2 L Tissue Culture-treated roller bottles (CorningTM)
50 ml/250 ml/5 L polycarbonate Erlenmeyer flasks (CorningTM)
Teflon/glass Dounce homogenizers (Cambridge Glassblowing Limited)
Miracloth (Merck Millipore Calbiochem, catalog number: 475855)
SW40 tubes for mitochondria gradient (Beckman, catalog number: 344060)
Polycarbonate bottle with cap assembly for type 70Ti rotor, for cushion (Beckman, catalog number: 355618)
TLS-55 tubes for mitoribosome gradient (Beckman, catalog number: 347356)
Dulbecco’s Modified Eagle Medium (GibcoTM, Life technologies)
Tetracycline-free fetal bovine serum (GibcoTM, Life technologies)
Freestyle 293 Expression medium (GibcoTM, Life technologies)
Dnase (RNA-free) (HT Biotechnology, N401a)
n-Dodecyl β-D-maltoside (Anatrace, catalog number: D310)
18:1 Cardiolipin (TOCL; Avanti, catalog number: 710335C)
β, γ-Methyleneguanosine 5′-triphosphate sodium salt (GMPPCP; Sigma, catalog number: M3509)
Protease inhibitor (cOmpleteTM, Roche)
Rabbit anti-mS27 (Abcam, catalog number: ab67157)
Mouse anti-OXA1L (Abcam, catalog number: ab88975)
Goat anti-mouse Alexa Fluor Plus 800 (ThermoFisher, catalog number: A32730)
Goat anti-rabbit Alexa Fluor Plus 680 (ThermoFisher, catalog number: A32734)
MIB Buffer (Mitochondria isolation buffer, see Recipes)
SM4 Buffer (Sucrose/mannitol buffer, see Recipes)
MIBSM Buffer (MIB+SM4: Experimental buffer, see Recipes)
SEM Buffer (Sucrose EDTA MOPS buffer, see Recipes)
60%, 32%, 23%, and 15% Sucrose Gradient Buffer (see Recipes)
Lysis Buffer (see Recipes)
Sucrose Cushion Buffer (see Recipes)
Resuspension Buffer (see Recipes)
15-30% Linear Sucrose Gradient Buffer (see Recipes)
Equipment
Automated cell counter (Countess® II FL)
Avanti-J26 XP centrifuge (Beckman Coulter)
High speed centrifuge (Beckman Coulter, model: Avanti J-26XP)
Ultracentrifuge (Beckman Coulter, model: Optima L-80 XP)
Tabletop ultracentrifuge (Beckman Coulter, model: OptimaTM MAX)
Centrifuge (Eppendorf, model: 5430 R)
NanoDrop® 2000 (Thermo)
Odyssey® CLx Infrared Imaging System (LI-COR Biosciences)
Vitrobot Mark IV (Thermo Fisher Scientific)
Titan Krios microscope (Thermo Fisher Scientific FEI)
Procedure
文章信息
版权信息
© 2021 The Authors; exclusive licensee Bio-protocol LLC.
如何引用
Readers should cite both the Bio-protocol article and the original research article where this protocol was used:
- Yang, H. and Desai, N. (2021). Purification of Mitochondrial Ribosomes with the Translocase Oxa1L from HEK Cells. Bio-protocol 11(15): e4110. DOI: 10.21769/BioProtoc.4110.
- Desai, N., Yang, H., Chandrasekaran, V., Kazi, R., Minczuk, M. and Ramakrishnan, V. (2020). Elongational stalling activates mitoribosome-associated quality control. Science 370(6520): 1105-1110.
分类
生物化学 > 蛋白质 > 分离和纯化
生物物理学 > 显微技术 > 低温显微镜技术
您对这篇实验方法有问题吗?
在此处发布您的问题,我们将邀请本文作者来回答。同时,我们会将您的问题发布到Bio-protocol Exchange,以便寻求社区成员的帮助。