- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Delayed Spatial Win-shift Test on Radial Arm Maze



Published: Vol 6, Iss 23, Dec 5, 2016 DOI: 10.21769/BioProtoc.2053 Views: 14205
Reviewed by: Xi FengXiaoyu LiuMarina Allerborn
Original research article
The authors used this protocol in:

Jun 2016
Advertisement


Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics









Abstract
The radial arm maze (RAM) is used to assess reference and working memory in rodents. This task relies on the rodent’s ability to orientate itself in the maze using extra-maze visual cues. This test can be used to investigate whether a rodent’s cognition is improved or impaired under a variety of experimental conditions. Here, we describe one way to test spatial working and reference memory. This delayed spatial win-shift (DSWS) procedure on the RAM was adapted from Packard and White (1990). The win-shift component of the test refers to the alternation of baiting, or rewarding, arms during the trial and test phase. The rodent is required to hold spatial information both within the task and across a delay to obtain the food-pellet reward (Taylor et al., 2003b). This task measures the incidence and type of memory errors made by the rodent both in the training and test phases of the learning task. A working memory error (re-entry of an arm that has been baited) can occur in both phases of the task, whilst a reference memory error (entry into an arm that has been baited during the training phase and is no longer baited) can only occur during the test phase.
Keywords: Delayed spatial win-shift (DSWS)Background
The radial arm maze (RAM) can be used to examine the effects of hippocampal and prelimbic cortex (PLC) damage, ageing, as well as a variety of pharmacological agents (Wenk, 2001; Taylor et al., 2003b; Floresco et al., 1997; Vann et al., 2003). The hippocampus is widely accepted to be involved in both spatial working and reference memory. Lesions to the hippocampus in rodents have shown impairments in the ability to perform memory tasks, including the RAM, involving spatial navigation (O'Keefe and Nadel, 1978; Morris et al., 1982). The PLC region of the rat prefrontal cortex, the approximate equivalent of primate dorsolateral region of the prefrontal cortex (Groenewegen, 1988), is also involved in spatial working memory (Robbins, 1990). Taylor et al. have demonstrated that rodents with lesions to the PLC make more spatial reference and memory errors compared to controls in the delayed spatial win-shift (DSWS) procedure on the radial maze (Taylor et al., 2003b). The traditional RAM studies an animal’s explorative behaviour during the task, particularly investigating working memory (Seamans et al., 1995). The adaptation of the task to include the DSWS element is a well-established procedure in the literature. This technique investigates the rats’ ability to retain spatial information both within the task and across a delay (Taylor et al., 2003b; Lapish et al., 2008; De Luca et al., 2016).
Materials and Reagents
- Paper towels for cleaning the maze with ethanol
- Rats: Our experiments were conducted using 10-week old male Wistar rats but other rodents can be used. If using females, estrous cycle stage should be taken into consideration since spatial reference memory is attenuated during the pro-estrous phase of the cycle (Bowman et al., 2001; Pompili et al., 2010). If using young rats or adult mice, a mouse RAM of smaller dimensions should be used, see below.
Notes:- The rats are housed under normal controlled laboratory conditions, in weight-matched pairs, under a 12 h dark/light cycle, with ad libitum access to food and tap water prior to the task. The testing protocol should be undertaken during the light phase of the 12 h light cycle between 0700 and 1900 h. Circadian rhythm does not need to be accounted for throughout the experiment as the rodents undergo bi-daily sessions across a large part of the 12 h light phase. To ensure there are no time biases between the groups, spread the testing of control and treated rodents throughout the day.
- During the testing protocol, access to food should be restricted to 80% of the rat’s usual food intake to encourage food-seeking behaviour in the maze. Adult (10-week old) male Wistar rats have an approximate daily food intake between 15-20 g during the 12 h dark phase and around 7 g during the 12 h light phase. Adult female Wistar rats have an approximate daily food intake between 10-15 g during the 12 h dark phase and around 5 g during the 12 h light phase (Stefanidis and Spencer, 2012).
- Prior to commencement and throughout the experiment, normal controlled laboratory light settings should be in place. The maximum allowable light intensity is equivalent to 300 lux at one meter height. There is no additional light source during the experimental protocol to alter the locomotor activity of the rodent.
- The rats are housed under normal controlled laboratory conditions, in weight-matched pairs, under a 12 h dark/light cycle, with ad libitum access to food and tap water prior to the task. The testing protocol should be undertaken during the light phase of the 12 h light cycle between 0700 and 1900 h. Circadian rhythm does not need to be accounted for throughout the experiment as the rodents undergo bi-daily sessions across a large part of the 12 h light phase. To ensure there are no time biases between the groups, spread the testing of control and treated rodents throughout the day.
- Ethanol, 70% (v/v), diluted in distilled water
- Standard chow grain pellets (45 mg) (Bio-Serv, USA)
Equipment
- RAM
The task is carried out in an eight-arm radial maze (Lafayette Instrument Company, USA), consisting of an octagonal central platform (34 cm diameter) and eight equally-spaced radial arms (87 cm long, 10 cm wide). At the end of each arm is a food well (2 cm in diameter and 0.5 cm deep; Figure 1). At the entrance to each arm is a clear Perspex door that controlled access in and out of the central area. Each door is controlled by a computerized control system (Lafayette) enabling the experimenter to regulate access to the arms. Salient visual cues of different geometric shapes and contrasting colours are placed around the maze on the walls of the room.
Note: If using young rats or adult mice, a mouse RAM of smaller dimensions should be used. The central platform should be approximately 22 cm in diameter with arms 25 cm long, 6 cm wide and 6 cm high that is transparent to enable the mice to see extra-maze visual cues (Crusio and Schwegler, 2005). The mouse RAM, should be placed on the floor to avoid elevation-induced anxiety (Crusio and Schwegler, 2005). - Digital video camcorder. In this experiment a Canon Legria FS200 was used. However, any camcorder can be used.
- Tripod. The video camcorder is attached to a tripod or to the ceiling to allow recording of the entire maze.

Figure 1. Radial Arm Maze (Lafayette Instrument Company) with automated Perspex doors and spatial cues placed around the maze. The camcorder is placed on the tripod facing downwards (approximately 45° to the maze). The camcorder can also be attached to the ceiling directly above the maze.
Procedure
- 2-day habituation
- During habituation, all eight arms are opened and the rodents are placed in the center arena facing away from the investigator (to give consistent introduction to the spatial cues) and allowed to freely explore the maze. Three standard chow grain pellets (45 mg) are placed along each arm to encourage exploration. Rodents are habituated to the maze for two consecutive days, with two 10 min sessions 4 h apart (Figure 2).
- After the final habituation session, the rodents are returned to their home cages and provided with approximately 20 grains of standard chow food pellets per animal.
- The arena is cleaned with 70% ethanol after each animal (and session) and allowed to dry thoroughly before the next rat is tested.
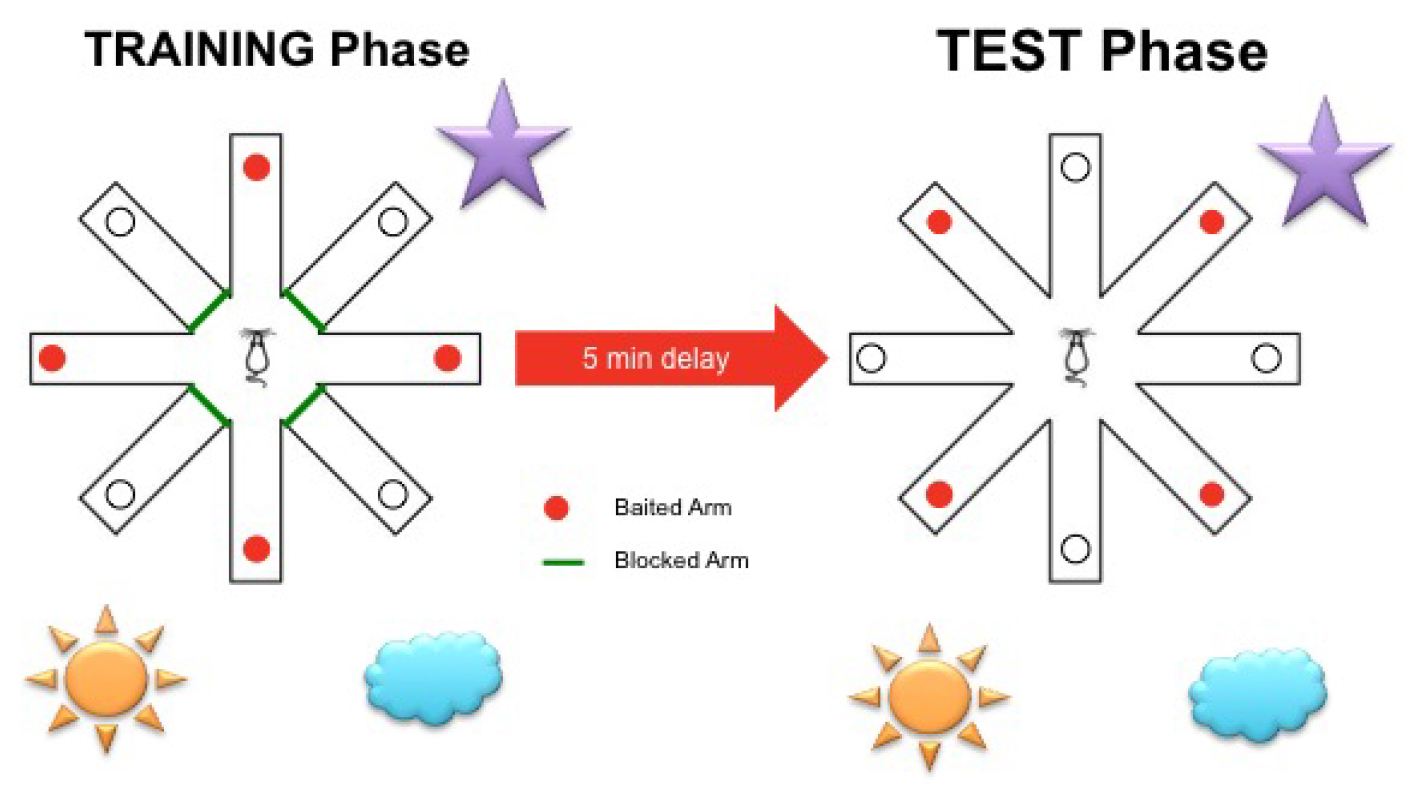
Figure 2. Delayed win-shift radial arm maze schematic representation with spatial cues placed around the maze (spatial cues in this figure are for illustrative purposes and were not the actual cues used). During the training phase, rodents are allowed to explore four pseudo-randomly selected arms baited with grain pellets whilst the other four arms are blocked. After a 5 min inter-trial-interval spent in their home cages, rodents are placed back into the maze with all arms accessible for the test phase. Previously blocked arms are then baited with grain pellets. - During habituation, all eight arms are opened and the rodents are placed in the center arena facing away from the investigator (to give consistent introduction to the spatial cues) and allowed to freely explore the maze. Three standard chow grain pellets (45 mg) are placed along each arm to encourage exploration. Rodents are habituated to the maze for two consecutive days, with two 10 min sessions 4 h apart (Figure 2).
- 13-day (26 trials) experimental phase
Note: The duration of the experimental phase is dependent on the control group making a mean of no more than one error in the test phase. Therefore this experiment may go for less or longer than 13 days (26 trials).- At the completion of the habituation phase, rodents are placed back into their home cages until the bi-daily training and test phases the following day. On day three the rodents undergo bi-daily sessions consisting of a 5 min training phase, a 5 min inter-trial interval (ITI) where the rodent is returned to its home cage, and a 5 min test phase. The bi-daily sessions are 5 h apart (Figure 3).
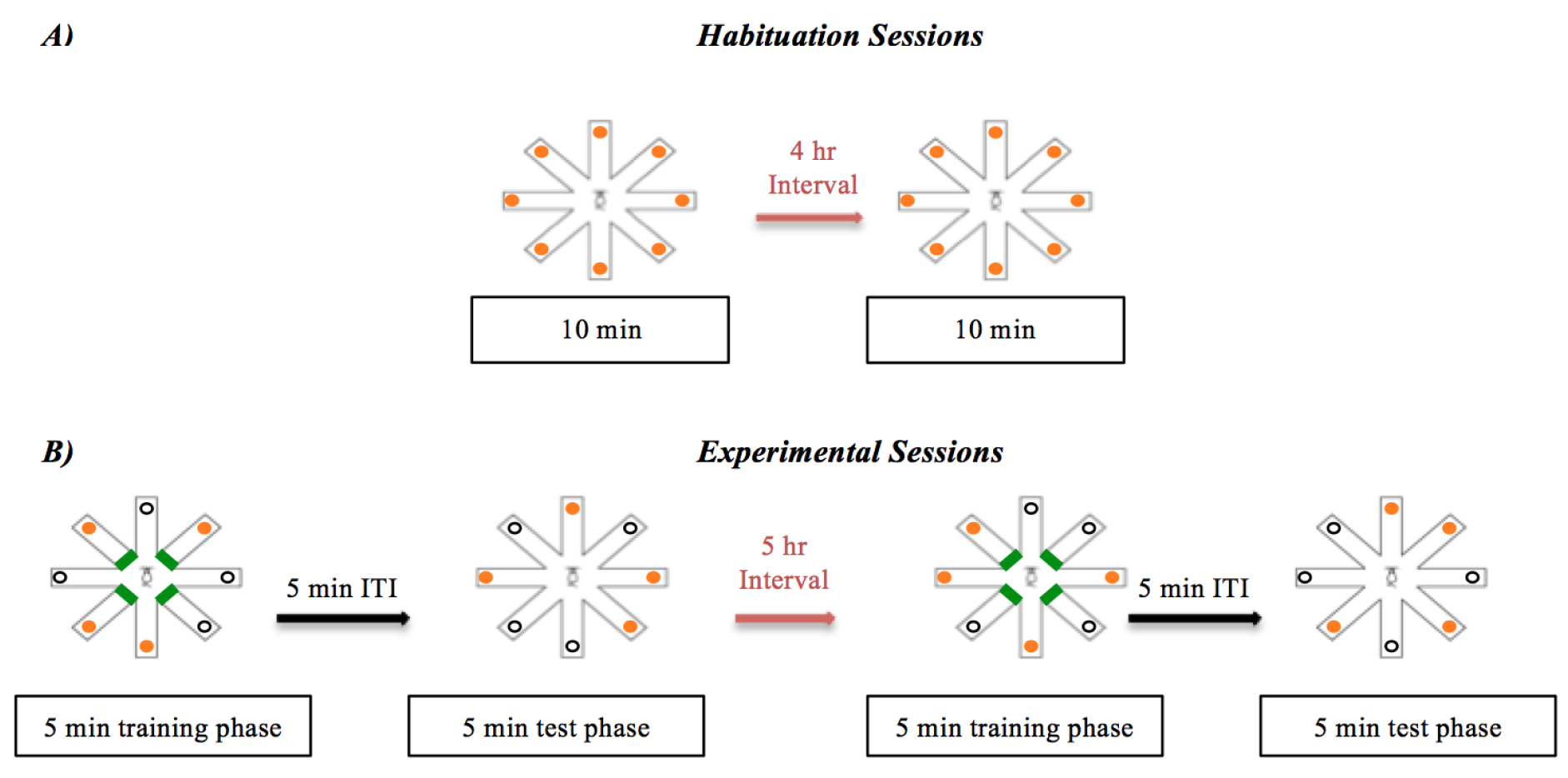
Figure 3. Timeline of the bi-daily habituation, training and test phases. A). Habituation sessions consist of two 10 min sessions with a 4 h interval. All eight arms are open and baited. B). Experimental bi-daily sessions. Each session includes a 5 min training phase, a 5 min inter-trial interval (ITI) and a 5 min test phase. - Throughout the experiment, access to standard chow food can be restricted to approximately 80% of the rodents’ normal food intake as recorded at the commencement of the experiment to encourage explorative behaviour during testing (Halagappa et al., 2007). Body weight is measured three times a week to ensure rodents do not fall below 80% free-feeding weight. If this does occur the amount of daily food intake can be increased. An alternative strategy is also to impose food deprivation during the dark cycle (Olton and Schlosberg, 1978).
- During the 5 min training phase, four of the eight arms are blocked. These are pseudo-randomly chosen (daily by the investigator) with no more than two adjacent arms being blocked at a time such that the combination of arms 1, 3, 4, 5 is excluded because 3, 4 and 5 are adjacent arms but 1, 2, 5, 7 is a valid combination because only 1 and 2 are adjacent (Floresco et al., 1997; Taylor et al., 2003b; Furgerson et al., 2014). The four unblocked arms are baited with three food pellets in each food well. During this phase of the experiment, the rodent is allowed to explore the maze for 5 min or until it has explored all of the open baited arms and retrieve the grain pellet rewards from all of them.
- The training phase can be video-recorded and scored using automated tracking programs such as EthovisionXT or can be manually scored in real time. An arm entry is recorded when the rodent moves all four paws off the central platform into the arm. Working errors (re-entry of an arm that had been baited and visited) can be recorded during this training phase.
- At completion of the training phase, rodents are returned to their home cage for 5 min. During this time, the maze is wiped down with 70% ethanol, allowed to dry completely and all eight arms opened. The previously blocked arms are baited with food pellets with the previously open arms left empty.
- After the ITI, rodents are placed back into the center of the maze and are allowed to explore all eight open arms for 5 min. This test phase can be video-recorded for automated scoring on camera or can be manually scored in real time. Two types of errors are recorded: working error (re-entry of an arm that had been baited and visited) and reference error (entry into an arm that had been baited during the training phase). The experiment is continued bi-daily until the control group makes a mean of no more than one error in the test phase (criterion), achieved after 26 trials in De Luca et al., 2016.
Troubleshooting: - If the rodent does not explore the maze during the habituation phase, a longer duration or additional sessions can be performed across the entire cohort before moving on to the test phase. The investigator may increase the amount of standard chow food pellets provided during the sessions as a greater incentive to explore.
- Rodents may attempt to jump from the arm they are exploring to an adjacent arm over the wall. To deter this from occurring the investigator should physically remove the rodent from the arm and place it back into the center of the maze facing away from the investigator.
- At the completion of the habituation phase, rodents are placed back into their home cages until the bi-daily training and test phases the following day. On day three the rodents undergo bi-daily sessions consisting of a 5 min training phase, a 5 min inter-trial interval (ITI) where the rodent is returned to its home cage, and a 5 min test phase. The bi-daily sessions are 5 h apart (Figure 3).
Data analysis
- To analyse the DSWS RAM, the training and test phase data and the type of memory error, either spatial working or reference error, are analysed separately. The training phase can only investigate spatial working memory errors, while the test phase can investigate both spatial working and reference memory errors.
- The mean of the number of errors from four consecutive trials is taken and formed into bins (Brown and Giumetti, 2006). For example if the rats reached criterion after 14 days (or 28 trials), the analysis would include seven data points (with each data point containing 4 trials). Example data are given in Figure 4.
- The data are analysed using two-way analyses of variance (ANOVA)s with treatment and block as between factors, followed by Tukey’s post hoc tests as described (De Luca et al., 2016). We assume statistical significance when P ≤ 0.05.
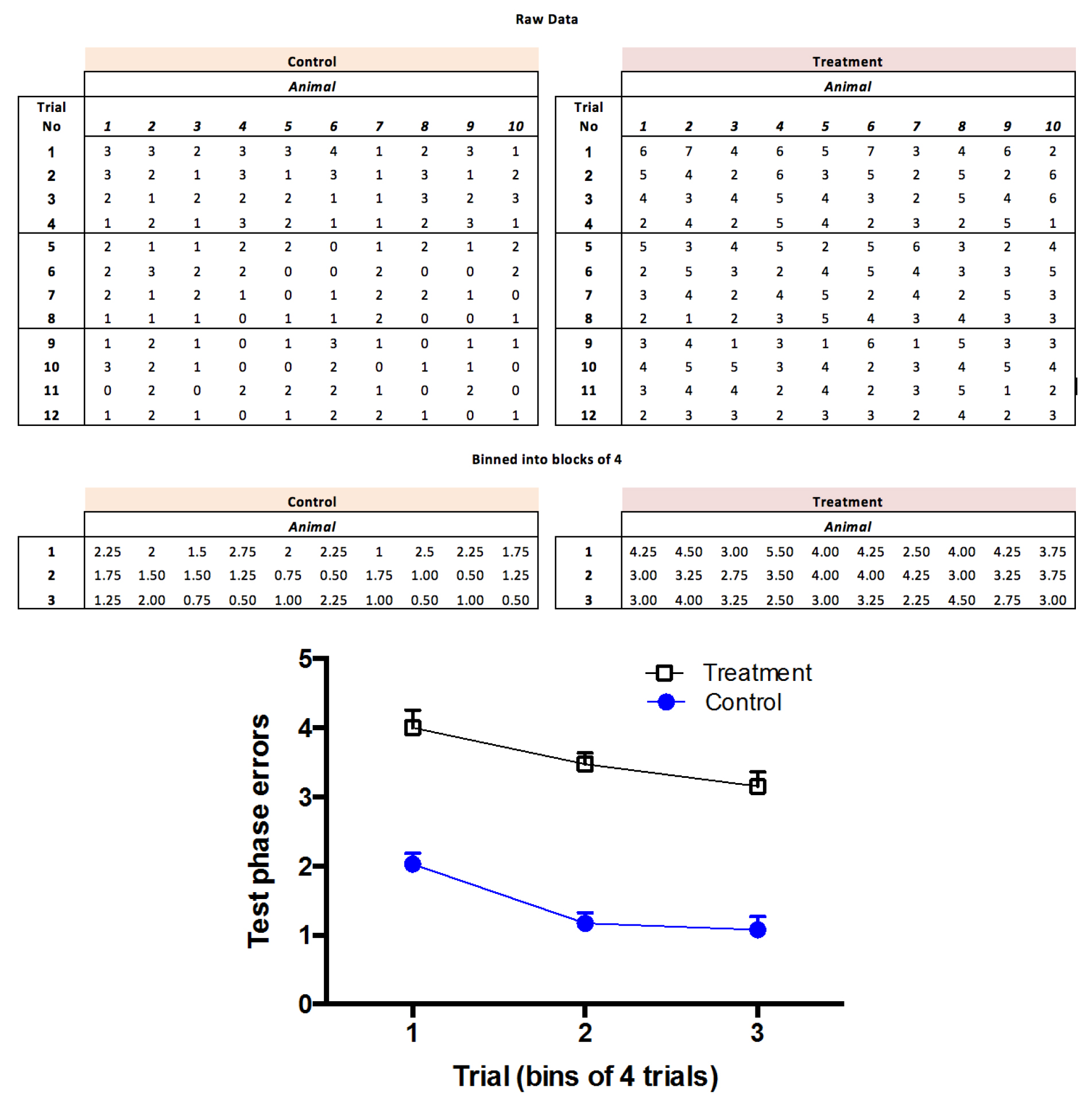
Figure 4. Example data showing incorrect arm entries during the test phase of the delayed win-shift radial arm maze. The numbers of incorrect entries from four consecutive test trials are binned together as a mean. The data are then analysed using two-way analyses of variance (ANOVA)s with treatment and block as between factors, followed by post hoc tests (De Luca et al., 2016).
Notes
- Statistical analysis can be performed on other measures using the DSWS procedure. The investigator can analyse the total number of errors made, the number of correct choices made before the first error, latency to reach the first arm from the central platform as well as the time it takes to complete the test (Taylor et al., 2003b).
- Spatial working and reference memory errors can often be described using other terms. Spatial working error (re-entry of an arm that had been baited and visited) is often referred to as within-phase errors and reference error (entry into an arm that had been baited during the training phase) as across-phase errors (Taylor et al., 2003b; Richter et al., 2013).
Acknowledgments
This behavioural procedure was adapted from previously published studies RAM (Packard and White, 1990; Taylor et al., 2003b; Taylor et al., 2003a; Floresco et al., 1997) and was performed by our group as described (De Luca et al., 2016). This work was supported by a Discovery Project Grant from the Australian Research Council (ARC) to SJ Spencer (DP130100508).
References
- Bowman, R. E., Zrull, M. C. and Luine, V. N. (2001). Chronic restraint stress enhances radial arm maze performance in female rats. Brain Res 904(2): 279-289.
- Brown, M. F. and Giumetti, G. W. (2006). Spatial pattern learning in the radial arm maze. Learn Behav 34(1): 102-108.
- Crusio, W. E. and Schwegler, H. (2005). Learning spatial orientation tasks in the radial-maze and structural variation in the hippocampus in inbred mice. Behav Brain Funct 1(1): 3.
- De Luca, S. N., Ziko, I., Sominsky, L., Nguyen, J. C., Dinan, T., Miller, A. A., Jenkins, T. A. and Spencer, S. J. (2016). Early life overfeeding impairs spatial memory performance by reducing microglial sensitivity to learning. J Neuroinflammation 13(1): 112.
- Floresco, S. B., Seamans, J. K. and Phillips, A. G. (1997). Selective roles for hippocampal, prefrontal cortical, and ventral striatal circuits in radial-arm maze tasks with or without a delay. J Neurosci 17(5): 1880-1890.
- Furgerson, M., Clark, J. K., Crystal, J. D., Wagner, J. J., Fechheimer, M. and Furukawa, R. (2014). Hirano body expression impairs spatial working memory in a novel mouse model. Acta Neuropathol Commun 2: 131.
- Groenewegen, H. J. (1988). Organization of the afferent connections of the mediodorsal thalamic nucleus in the rat, related to the mediodorsal-prefrontal topography. Neuroscience 24(2): 379-431.
- Halagappa, V. K., Guo, Z., Pearson, M., Matsuoka, Y., Cutler, R. G., Laferla, F. M. and Mattson, M. P. (2007). Intermittent fasting and caloric restriction ameliorate age-related behavioral deficits in the triple-transgenic mouse model of Alzheimer's disease. Neurobiol Dis 26(1): 212-220.
- Lapish, C. C., Durstewitz, D., Chandler, L. J. and Seamans, J. K. (2008). Successful choice behavior is associated with distinct and coherent network states in anterior cingulate cortex. Proc Natl Acad Sci U S A 105(33): 11963-11968.
- Morris, R. G., Garrud, P., Rawlins, J. N. and O'Keefe, J. (1982). Place navigation impaired in rats with hippocampal lesions. Nature 297(5868): 681-683.
- O'Keefe, J. and Nadel, L. (1978). The Hippocampus as a Cognitive Map. Clarendon Press.
- Olton, D. S. and Schlosberg, P. (1978). Food-searching strategies in young rats: Win-shift predominates over win-stay. J Comp Physiol Psychol 92(4): 609.
- Packard, M. G. and White, N. M. (1990). Lesions of the caudate nucleus selectively impair "reference memory" acquisition in the radial maze. Behav Neural Biol 53(1): 39-50.
- Pompili, A., Tomaz, C., Arnone, B., Tavares, M. C. and Gasbarri, A. (2010). Working and reference memory across the estrous cycle of rat: a long-term study in gonadally intact females. Behav Brain Res 213(1): 10-18.
- Richter, S. H., Zeuch, B., Lankisch, K., Gass, P., Durstewitz, D. and Vollmayr, B. (2013). Where have I been? Where should I go? Spatial working memory on a radial arm maze in a rat model of depression. PloS one 8(4): e62458.
- Robbins, T. W. (1990). The prefrontal cortex, 2nd edn. In J. M. Fuster (Ed.). International Journal of Geriatric Psychiatry. Raven, pp:348-348.
- Seamans, J. K., Floresco, S. B. and Phillips, A. G. (1995). Functional differences between the prelimbic and anterior cingulate regions of the rat prefrontal cortex. Behav Neurosci 109(6): 1063-1073.
- Stefanidis, A. and Spencer, S. J. (2012). Effects of neonatal overfeeding on juvenile and adult feeding and energy expenditure in the rat. PLoS One 7(12): e52130.
- Taylor, C. L., Latimer, M. P. and Winn, P. (2003a). Impaired delayed spatial win-shift behaviour on the eight arm radial maze following excitotoxic lesions of the medial prefrontal cortex in the rat. Behav Brain Res 147(1-2): 107-114.
- Taylor, C. L., Latimer, M. P. and Winn, P. (2003b). Impaired delayed spatial win-shift behaviour on the eight arm radial maze following excitotoxic lesions of the medial prefrontal cortex in the rat. Behav Brain Res 147(1-2): 107-114.
- Vann, S. D., Kristina Wilton, L. A., Muir, J. L. and Aggleton, J. P. (2003). Testing the importance of the caudal retrosplenial cortex for spatial memory in rats. Behav Brain Res 140(1-2): 107-118.
- Walton, W. E. (1933). Color vision and color preference in the albino rat. II. The experiments and results. J Comp Psychol 15(3): 373.
- Wenk, G. L. (2004). Assessment of spatial memory using the radial arm maze and Morris water maze. Current protocols in neuroscience 8-5.
Article Information
Copyright
© 2016 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
De Luca, S. N., Sominsky, L. and Spencer, S. J. (2016). Delayed Spatial Win-shift Test on Radial Arm Maze. Bio-protocol 6(23): e2053. DOI: 10.21769/BioProtoc.2053.
Category
Neuroscience > Behavioral neuroscience > Animal model > Mouse
Neuroscience > Behavioral neuroscience > Cognition
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.