- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

Cytohistological Analyses of Mega-sporogenesis and Gametogenesis in Ovules of Limonium spp.

Published: Vol 6, Iss 21, Nov 5, 2016 DOI: 10.21769/BioProtoc.1983 Views: 7698
Reviewed by: Marisa RosaAleksandr GavrinAnonymous reviewer(s)
Original research article
The authors used this protocol in:

Jan 2016
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics







Abstract
Limonium spp. are known to have sexual and apomixis (asexual reproduction through seeds) reproductive modes. Here, we present dissection protocol developed for ovules of Limonium spp. using differential interference contrast (DIC) microscopy. This protocol permits better handling of ovules and offers certain advantages over earlier techniques particularly in larger ovules. This method also enables observation of meiosis and embryo sac development in intact ovules, and the ready detection of events distinguishing sexual and apomictic development.
Background
To describe the events that occur during ovule development it is necessary to cytologically examine ovules. This study can involve microscopic observation of paraffin- or resin-embedded, sectioned material, or cleared organs. The first cytological investigations into ovule and embryo sac development in sexual and apomictic Limonium species were published in the pioneer works of D’Amato (1940; 1949). In these works, flowers were fixed using the Karpechenko’s method, embedded in paraffin, sectioned and stained with Heidenhain’s iron haematoxylin, which stains chromatin and chromosomes in the cell nuclei. Flower buds sectioning using these methods can result in preparations with poor quality, due to partial disruption structural integrity of individual cells. A more facile alternative is clearing formalin:acetic acid:ethyl alcohol fixed organs and staining with pure Mayer’s hemalum (Wallis, 1957; Stelly et al., 1984). This technique requires much less time and labor, particularly for species which usually only form a small ovule within the ovary, which is the case of Limonium spp. However, in both small and large ovules chloral hydrate worked better than methyl salicylate as a clearing solution, because in this latter fluid ovules become quite fragile and difficult to handle during experiments. Our approach with an enzymatic digestion of ovules helps to reveal the central mass of tissue, the nucellus and the two integuments it covers, particularly in large ovules. Examples of meiotic and ameiotic ovules and embryo sacs cleared in chloral hydrate were observed under differential interference contrast optics.
Materials and Reagents
- Glass microscope slides (Belden, Hirschmann, catalog number: 8210101 )
- Diagnostic microscope multiwell slides (Thermo Fisher Scientific, Thermo ScientificTM, catalog number: 101432648 EPOXY )
- Glass coverslips (24 x 50 mm) (Belden, Hirschmann, catalog number: 8000119 )
- Teasing needle (BioQuip Products, catalog number: 4751 )
- Micropipette tips (2-20 μl, 20-200 μl, 100-1,000 μl) (Eppendorf)
- Six to eight months old Limonium plants in the flowering period
- Formaldehyde (36% aqueous solution) (VWR, catalog number: 20909.290 )
- Glacial acetic acid (Thermo Fisher Scientific, Fisher Scientific, catalog number: 10304980 )
- Distilled H2O
- Ethanol (absolute 99.5%) (Thermo Fisher Scientific, Fisher Scientific, catalog number: BP2818500 )
- Hematoxylin solution, Mayer’s (Sigma-Aldrich, catalog number: MHS16 )
- Chloral hydrate (Sigma-Aldrich, catalog number: 15307 )
- Cellulase (Sigma-Aldrich, catalog number: C1184 )
- Cellulase ‘Onozuka R-10’ (Serva, catalog number: 16419 )
- Pectinase (Sigma-Aldrich, catalog number: 6287 )
- Citric acid-1-hydrate (Sigma-Aldrich , catalog number: 33114 )
- Sodium citrate dihydrate (AppliChem, catalog number: 131655 )
- FAA solution (see Recipes)
- Dehydration solutions (see Recipes)
- Hydration solutions(see Recipes)
- 0.1% chloral hydrate (see Recipes)
- Enzymatic mixture (see Recipes)
- 1x EB (Enzyme buffer) (see Recipes)
Equipment
- Forceps with extra fine tips (Rubis forceps [BioQuip Products, catalog number: 4524 ])
- Glass watch glasses (O.D. 25 mm and 40 mm) (Sigma-Aldrich)
- Stereo microscope (Leica, model: WILD M3Z LEITZ )
- Fluorescence microscope (Carl Zeiss, model: Axioskop 2 )
- Differential interference contrast (DIC) optics
- Micropipettes (2-20 μl, 20-200 μl, 100-1,000 μl) (Eppendorf)
- AxioCam 289 MRc5 digital camera (Carl Zeiss)
Software
- Adobe Photoshop 5.0
Procedure
- Collect flower buds at different developmental stages ranging from 2 to 6 mm in length (Figure 1A, left – young stage; right – mature stage) using forceps. Flower buds are fixed in 1 ml FAA solution in a watch glass for 24 h at 4 °C. The fixed material may be stored in 1 ml 70% ethanol at -20 °C until use for 3 months. To this end, after fixation the flower buds should be dehydrated in a series of water/ethanol (v/v) solutions, 30%, 50%, 70%, shaking for 30 min each before storage.
Note: As the flowering season occurs during a short period of the year, storage can be useful to save flower buds for further use. - Dissect pistils from flower buds with forceps onto a microscope slide using a stereomicroscope. Make a sharp longitudinal incision in the ovary (Figure 1B, incision arrowed) with a needle to help subsequent ovule fixation in 1 ml FAA solution for 14-18 h, at 4 °C.
Note: Alternatively, big ovaries (> 5 mm) can be dissected directly in a drop (5 μl) of FAA, and then proceed to step 3.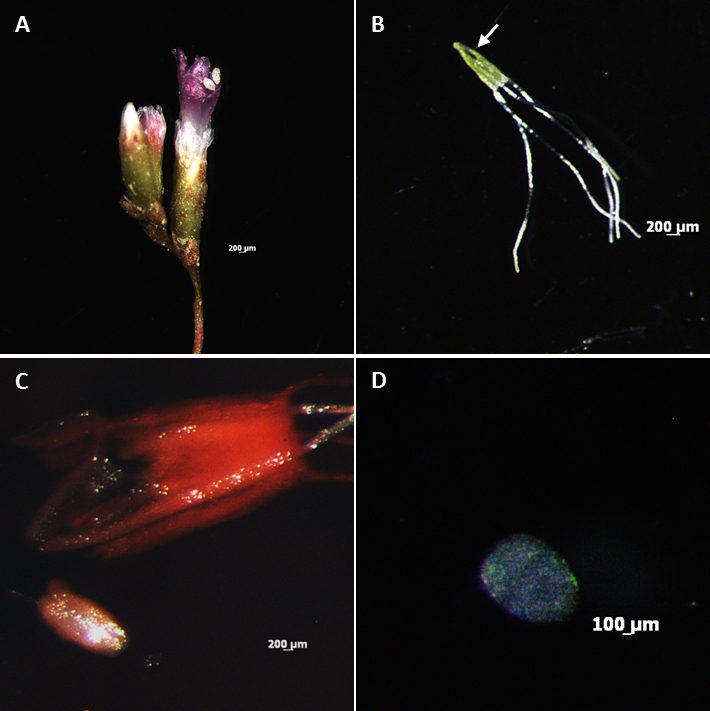
Figure 1. Dissection procedure and obtainment of digested ovules of Limonium spp. A. Flower buds at different developmental stages ranging from 2 to 6 mm (Figure 1A: left – young stage; right – mature stage). B. Pistil showing a longitudinal incision in the ovary (arrowed). C. Stained ovule immediately after dissection. D. Digested ovule mounted in a drop of chloral hydrate, in well of a multiwell slide.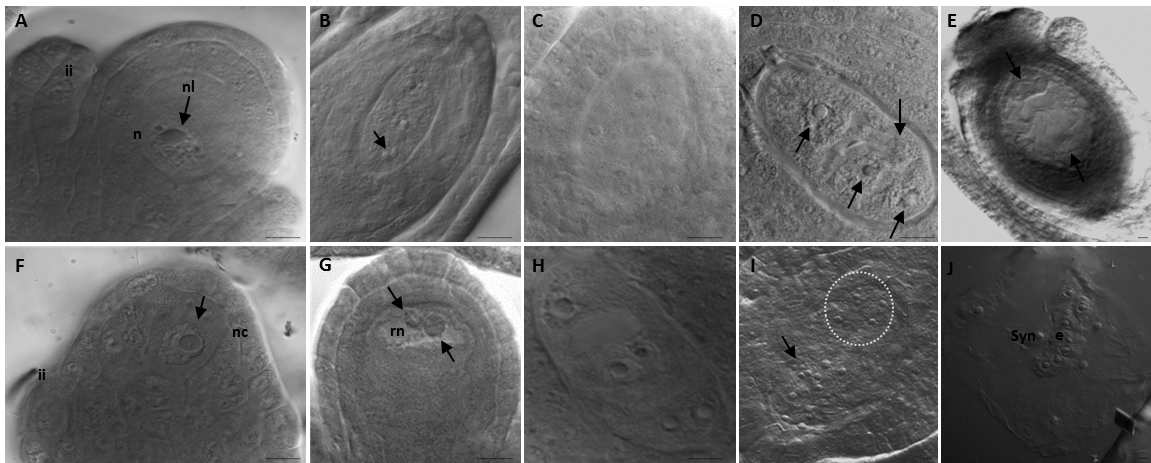
Figure 2. Examples of meiotic and ameiotic ovules and embryo sacs cleared in chloral hydrate and observed under differential interference contrast (DIC) optics. Figures A-E show sexual diploid (2n = 2x = 16) Limonium ovalifolium plants: accession numbers 2009I4SR (Sra. Remédios, Peniche), and 2009I1PL (Praia da Luz, Lagos), 2009I1PL (Praia da Luz, Lagos). Figures F-J were taken from apomictic tetraploid (2n = 4x = 35, 36) Limonium multiflorum plants: accession numbers 2009I4VF (Vale dos Frades, Lourinhã), 2009I9CR (Cabo Raso, Cascais), 2009I4VF (Vale dos Frades, Lourinhã). A. Megaspore mother cell at interphase with one nucleolus (arrowed). B. Coenomegaspore with four unreduced nuclei, each nucleus with two nucleoli (arrowed) (4-nucleated stage; tetrasporic development). C. Coenomegasporocyte at the beginning of gametogenesis with a cruciate nuclear arrangement. D. Megaspores in a bipolar 1 + 3 arrangement (arrowed) with the embryo sac showing extended vacuolization. E. Detail of an embryo sac of Adoxa-type with the egg cell and inferior polar nuclei arrowed. F. Megasporocyte within ovule showing one nucleolus (arrowed). G. Restitution nuclei after first meiotic division (meiotic diplospory). Nucleoli with distinct dimensions within the same nucleus are arrowed. H. Coenomegasporocyte at the beginning of gametogenesis. I. Detail of the egg cell apparatus (encircled by dashed lines) in a mature embryo sac; antipodal cells towards the chalazal pole are arrowed. J. Parthenogenetic embryo formation within a mature sac. Bars = 10 μm.
Note: Each image is oriented so that the micropyle is towards the top of the page and the chalaza is towards the bottom. Figures abbreviations: e – embryo; ii – integuments; n – nucleus; nl – nucleolus; nc – nucellus; rn – restitution nucleus; Syn – synergid. - Afterwards, hydrate pistils in watch glasses in a series of progressive 1 ml water/ethanol solutions, 50%, 75% and 100%, shaking for 30 min each.
- Stain pistils with 1 ml pure Hemalum Mayer’s solution for 24-48 h in a watch glass, and then place them in 1 ml 0.1% chloral hydrate in a new watch glass during 1-2 h, at room temperature.
Note: If the ovaries are small (< 3 mm) 24 h in the staining solution are required; for bigger ones 48 h may be needed. - Thereafter, dissect ovules from ovaries (Figure 1C) onto a glass slide under a stereomicroscope. Then, place them in a watch glass for digestion with 500 μl of an enzyme mixture 2% (w/v) cellulase, 2% (w/v) cellulase ‘Onozuka R-10’, and 2% (v/v) pectinase in 1x EB, for 10 (young stages) to 20 min (mature stages) at 37 °C, inside a box on a paper humidified with distilled water.
- Remove the enzyme mixture by a micropipette and wash ovules in a watch glass, in 1 ml 1x EB solution for 3-5 min, at room temperature.
- Finally, mount ovules in multiwell slides in a drop (5 μl) of chloral hydrate, under a coverslip (Figure 1D).
- When the procedure is completed, cytohistological observations are made using a fluorescence microscope with DIC optics using 20x and 100x objectives (Figure 2).
- Images are collected with a digital camera and if necessary further processed using Adobe Photoshop 5.0.
Data analysis
Female gametophytes of sexual and/or apomictic origin were determined in accessions from the ISA collections, and cytohistological observations were made under 20x and 100x magnification. For each species the development pattern ovules during megasporogenesis and gametogenesis were examined, and the number and frequency of tetrasporic versus diplosporous embryo sacs were evaluated (c. 700). In sexual species tetrasporic embryo sacs of Gagea-ova, Adoxa- and Drusa-types were found while apomictic species only produced diplosporous meiotically-unreduced eggs of the Rudbeckia-type. Details on numerical analysis of these ovules are published in Róis et al. (2016).
Recipes
- FAA solution
Formaldehyde-acetic acid-ethyl alcohol in proportions of 1:1:18 (38.5% formaldehyde, 5% acetic acid, 70% ethanol)
For 100 ml mix:
5 ml of 38.5% formaldehyde
5 ml of glacial acetic acid
90 ml of 70% ethanol
Note: Use always a fresh solution. Do not store it. - Dehydration solutions
30% EtOH-70% H2O
50% EtOH-50% H2O
70% EtOH-30% H2O
Note: Make fresh solutions every three months. - Hydration solutions
50% EtOH-50% H2O
25% EtOH-75% H2O
100% H2O
Note: Make fresh solutions every three months. - 0.1% chloral hydrate
For 50 ml mix:
50 mg of chloral hydrate
50 ml of distilled water
Notes:
a.Make a fresh solution every three months.
b.Chloral hydrate is toxic if swallowed. Can be irritating to eyes and skin.
c.Store in a cool, dry, well ventilated place, in securely closed original container. - Enzymatic mixture
For 20 ml:
606 µl pectinase
0.12 g cellulase ‘Onozuka R10’
1.10 g cellulase Sigma C-1184
Add 20 ml 1x PBS
Note: Store 500 µl aliquots at -20 °C. - 1x EB (enzyme buffer)
For 100 ml mix:
40 ml of 1 M citric acid-1-hydrate
60 ml of 1 M sodium citrate dihydrate
pH 4.8
Note: 1x EB buffer should be kept at 4 °C for 1-2 weeks. For longer periods of time, buffer should be stored at -20 °C.
Acknowledgments
We thank Generosa Teixeira (Faculdade de Farmácia, University of Lisbon) for useful discussions with her on methods. FUNDAÇÃO PARA A CIÊNCIA E TECNOLOGIA provided financial support for this work under project AGR/04129 (LEAF).
References
- D’Amato, F. (1940). Contributo all’embriologia delle Plumbaginaceae. Nuovo Giornale Botanico Italiano 47(2): 349-382.
- D’Amato, F. (1949). Triploidia e apomissia in Statice oleaefolia Scop. var. confusa Godr. Caryologia 2(1): 71-84.
- Róis, A. S., Sadio, F., Paulo, O. S., Teixeira, G., Paes, A. P., Espirito-Santo, D., Sharbel, T. F. and Caperta, A. D. (2016). Phylogeography and modes of reproduction in diploid and tetraploid halophytes of Limonium species (Plumbaginaceae): evidence for a pattern of geographical parthenogenesis. Ann Bot 117(1): 37-50.
- Stelly, D. M., Peloquin, S. J., Palmer, R. G. and Crane, C. F. (1984). Mayer’s hemalum-methyl salicylate: a stain-clearing technique for observations within whole ovules. Stain Technol 59(3): 155-161.
- Wallis T.E. 1957. Analytical microscopy. London: J & A Churchill.
Article Information
Copyright
© 2016 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Róis, A. S. and Caperta, A. D. (2016). Cytohistological Analyses of Mega-sporogenesis and Gametogenesis in Ovules of Limonium spp.. Bio-protocol 6(21): e1983. DOI: 10.21769/BioProtoc.1983.
Category
Plant Science > Plant cell biology > Cell imaging
Cell Biology > Cell imaging > Fixed-cell imaging
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.