- Protocols

- Articles and Issues
- For Authors
- About
- Become a Reviewer

A Method to Analyze Local and Systemic Effects of Environmental Stimuli on Root Development in Plants


Published: Vol 6, Iss 17, Sep 5, 2016 DOI: 10.21769/BioProtoc.1923 Views: 10952
Reviewed by: Renate WeizbauerAlizée MalnoeHonghong Wu
Original research article
The authors used this protocol in:

Jun 2014
Advertisement



Protocol Collections
Comprehensive collections of detailed, peer-reviewed protocols focusing on specific topics







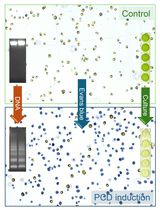

Abstract
Root development in vascular plants is innately tied to the environment. However, relatively little attention has been paid toward understanding the spatial scales at which the root perceives and responds to external stimuli. While some environmental signals elicit global responses that affect root system architecture, others may have more localized effects. We have observed that various developmental processes can be induced or suppressed along the circumference of the main root depending on local contact with available water in a process termed hydropatterning (Bao et al., 2014). Our studies of hydropatterning indicate that the root can detect and respond to certain external stimuli at the resolution of the diameter of a single organ. In order to characterize developmental patterning at this spatial scale, we developed a procedure to vary environmental inputs across the circumferential axis of the root in vitro using agar media. Roots are grown between two blocks of agar media in a “sandwich”. Local environmental conditions can be varied depending on the composition of the media on either side. Stimuli that act locally can be distinguished from those that act systemically based on the developmental response of the root. Here we describe the overall method and provide an example of how it can be used to analyze lateral root patterning in Zea mays (maize) in response to an external water potential gradient. We also discuss how the method can be used more broadly for other plant species and environmental treatments.
Keywords: Plant-environment interactionsMaterials and Reagents
- Germination paper (76 lb heavy weight seed germination paper) (Anchor Paper Company)
- 120 x 120 x 17 mm square plastic Petri plates (USA Scientific, catalog number: 5668-8102 )
- Aluminum foil
- Tri-fold paper towels (von Drehle Corporation, catalog number: 548-W )
- Silicone rubber (food-grade, high-temperature, 1/32” thickness, 50A medium hardness) (McMaster-Carr, catalog number: 86045K131 )
- Razor blades
- 50 ml conical tubes
- Filter paper, Grade 40 (Sigma-Aldrich, Whatman®, catalog number: Z241261 )
- Parafilm (VWR International, Bemis, catalog number: PM999 )
- Distilled water
- MS salts (Caisson Laboratories, catalog number: MSP01-50LT )
- MES hydrate (Sigma-Aldrich, catalog number: M2933 )
- Potassium hydroxide (1 M aqueous solution)
- Agar (BD, DifcoTM, catalog number: 214530 )
- Mannitol (PhytoTechnology Laboratories, catalog number: M562 )
- Maize kernels (B73 inbred line)
- Bleach (8.25% sodium hypochlorite solution)
- Tween 20 (Sigma-Aldrich, catalog number: P2287 )
- 70% (v/v) ethanol in water
- Superglue (Krazy Glue, catalog number: KG582 )
- Control agar media (see Recipes)
- Treatment agar media (see Recipes)
- Maize kernel sterilization solution (see Recipes)
Equipment
- Glass bottles (500 ml capacity or greater)
- pH meter
- Autoclave
- Laminar flow hood
- 4 °C cold room or refrigerator
- Glass beakers (500 ml capacity or greater)
- Hot water bath or hot plate
- Thermometer
- Timer
- Forceps
- Plant growth chamber (Percival, model number: CU41L4 )
- Permanent marker
- Microwave
- Ruler
- Computer equipped to run R or comparable statistical analysis software
Software
- R or comparable statistical analysis software (download link: https://cloud.r-project.org/)
Procedure
- Agar media preparation and sterilization of materials for seed plating and agar sandwich assembly
Note: These steps should be performed no later than one day following seed sterilization (see below). These supplies can be stored for weeks; preparing them well ahead of time is recommended.- Prepare control and treatment agar media
Note: Four possible agar sandwich conditions can be constructed given two media types (block side/plate side: control/control, treatment/control, control/treatment, and treatment/treatment – see step C7 for further information). 1 L (15 plates) of each media type is sufficient to grow 12 plants per condition. This assumes two plants are used per plate, and includes extra plates for cutting agar blocks (see step C7). Amounts of media necessary will vary depending on desired sample size and number of conditions. - Autoclave distilled water for 30 min at 121 °C on wet cycle. 500 ml autoclaved water is enough to sterilize ~100 seeds.
Note: To save time, autoclave distilled water and agar media in the same cycle. - Cut germination paper into 12 x 12 cm squares and wrap in aluminum foil. Two squares are sufficient to germinate 10-12 seeds.
- Fold paper towels along tri-fold creases and cut in half along the short axis. Wrap half-sheets in aluminum foil. Prepare one paper towel half-sheet for every 10-12 plants.
- Cut silicone rubber into 1.5 x 6 cm strips and wrap in aluminum foil. Cut two strips for each plant.
- Cut filter paper into ~4 x 4 cm squares and wrap in aluminum foil. Cut one square for each plant.
- Wrap 3-4 razor blades in aluminum foil.
- Autoclave germination paper squares, paper towel half-sheets, silicone rubber strips, filter paper squares, and razor blades at 121 °C for 20 min on dry cycle.
Note: It is not recommended to autoclave these items along with agar media and distilled water, which should be autoclaved on wet cycle.
- Prepare control and treatment agar media
- Sterilization and germination of maize kernels
- Soak seeds for 4 h in distilled water in 50 ml conical tubes.
Note: Use enough water to completely cover all of the kernels. For ease of handling, fill the tubes no higher than the 30 ml mark with kernels. - Decant water. Remove tip caps from the kernels using a razor blade, ensuring that the embryo and endosperm of each kernel are not damaged (Figure 1A). Place kernels back in the 50 ml conical tube.
Note: On occasion, the tip cap of the kernel may be too small or inconvenient to access without damaging other parts of the seed. In such cases, the excision step can be skipped. - Incubate kernels in warm water (55-57 °C) for 5 min. Ensure that kernels are placed directly in contact with warm water by pouring some into the 50 ml conical tube containing the kernels before placing the tube in the water bath.
Note: This can be done using a hot water bath, or by heating water in a glass beaker with a hot plate. Use of sterile water is not required at this point. - Decant warm water. Rinse seeds once with room-temperature water.
- Incubate kernels in maize kernel sterilization solution for 20 min. Shake the tube occasionally (once every few minutes).
- Conduct all following steps in a laminar flow hood. Spray down all working surfaces and materials with 70% ethanol before placing into the hood.
- Decant maize kernel sterilization solution. Waste solution can be collected in a beaker.
- Rinse kernels 5-7 times with autoclaved distilled water.
- Plate seeds on autoclaved germination paper
- Place two germination paper squares into a plastic Petri plate. Place a paper towel half-sheet on top of the germination paper in the center of the plate (Figure 1A).
- Add 20 ml autoclaved distilled water to the plate to saturate the paper. Prepare enough plates for the number of kernels sterilized (10-12 kernels per plate).
- Fold back the paper towel half-sheet. Arrange 10-12 kernels in a row along the paper towel. Orient the kernels so that the cut ends point away from the paper towel (Figure 1B).
- Fold the paper towel half-sheet over the kernels. Smooth down the paper towel against the germination paper to limit movement of the kernels (Figure 1C).
- Seal the plates closed with Parafilm.
- Place two germination paper squares into a plastic Petri plate. Place a paper towel half-sheet on top of the germination paper in the center of the plate (Figure 1A).
- Incubate plates vertically in a growth chamber for 2-3 days at 29 °C, 16 h light/8 h dark cycle. Orient the plates so that the cut ends of kernels are pointing toward gravity.

Figure 1. Preparation of maize kernels for germination on sterile germination paper in square plates. A. A maize kernel with the embryo, endosperm, and tip cap outlined in black, cyan, and red, respectively. B. Square Petri plate containing germination paper and paper towel after soaking in water. C. Sterilized maize kernels lined up on germination paper. Note that the cut ends of the kernels point away from the paper towel that has been folded back. D. Germination paper plate after sealing with Parafilm. The paper towel has been folded over the kernels and smoothed down against the germination paper to keep the kernels in place. E. Germinated seedlings two days after plating.
- Soak seeds for 4 h in distilled water in 50 ml conical tubes.
- Agar sandwich assembly
- Conduct all steps in a laminar flow hood. Spray down all working surfaces and equipment with 70% ethanol before placing into the hood. Use forceps to manipulate materials such as agar, filter paper, silicone rubber, etc.
- Remove the Parafilm from the germination paper plates and fold back the paper towels. Check that primary roots have emerged and grown to a length of ~1 cm (Figure 1D).
- Take a plate of control agar media and cut a ~4 x 9 cm rectangle near one edge of the plate using an autoclaved razor blade. Remove and discard the agar in the rectangle. Ensure that the resulting hole is completely free of residual agar. This plate will be used for the “plate side” of the agar sandwich (Figure 2A).
- Place two squares of filter paper below the rectangular hole in the agar plate (Figure 2B). Smooth the paper down against the agar to remove any bubbles.
- Place silicone rubber strips to the left and right of the filter paper squares. Position the strips so that they just overlap the filter paper (Figure 2C).
- Glue the rubber to the agar using a small amount (1-2 drops) of superglue (Figure 2D).
Note: Proceed carefully when positioning rubber strips, as superglue bonds very quickly and repositioning after the adhesive has set is challenging. Do not use too much superglue; although we rarely observe negative effects on the plant from superglue, root growth and development become strongly inhibited if the root comes into direct contact with either liquid or solid glue. - Take another agar plate and cut it into 9 equally-sized squares using an autoclaved razor blade. These will be used for the “block side” of the agar sandwiches.
Note: A printable template has been included in this protocol to assist with cutting agar blocks to the correct size (Figure 3). - Remove two blocks from the plate in step C7 and place a filter paper square on each of them. Cut away any overhanging filter paper using an autoclaved razor blade (Figure 2E). Place the blocks paper-side-down into an empty plate lid to assist in cutting the filter paper.
- Position two seedlings in the agar plate from step C6 such that the kernels sit in the rectangular hole and the primary root tips are on top of the filter paper (Figure 2F). Avoid touching the primary root or coleoptile directly with the forceps.
- Glue down the agar blocks over the primary roots of the seedlings.
- Put a few drops of superglue on the exposed sides of the silicone rubber in the agar plate with the seedlings (Figure 2G).
- Lay the agar blocks across the silicone rubber rectangles with the paper side facing the seedlings (Figure 2H). Line up the blocks with the filter paper squares in the agar plate.
- Press the blocks against the silicone rubber rectangles so that the glue adheres them in place.
- Put a few drops of superglue on the exposed sides of the silicone rubber in the agar plate with the seedlings (Figure 2G).
- Seal the agar sandwich plate with Parafilm.
- If a seedling comes out of the sandwich, perform either of the following:
- Reposition the seedling and slide the primary root further into the sandwich.
- Reposition the seedling so that the kernel is back in the rectangular hole in the plate. Cut a block of agar from an unused plate (either control or treatment media to match the media type on the plate-side of the seedling) and place it in a 50 ml conical tube. Heat the agar in a microwave until it melts (about 10-15 sec), and pipet the cooled molten agar solution around the kernel. After the agar solidifies, the seedling will be fixed in place.
- Reposition the seedling and slide the primary root further into the sandwich.
- Label the plate with the agar sandwich condition (denoted as “block side/plate side”, e.g., “control/control”).
- Mark the positions of the primary root tips on the back of the plate using a permanent marker. Make sure the mark is visible from the other side of the plate (Figure 2I).

Figure 2. Assembly of agar sandwiches. A. Plate-side agar media after removal of rectangular section of agar. B. Positioning of filter paper squares on plate-side agar media. C. Positioning of silicone rubber strips over filter paper squares. D. Applying superglue to one of the silicone rubber strips. Note that only a small volume of glue is required at each point of application. E. Agar blocks cut from a separate plate of media. Blocks are depicted sitting in an empty plate lid with filter paper squares applied. F. Maize seedlings in position in the plate containing the plate-side of the agar sandwich. G. Application of superglue to the silicone rubber strips before applying the agar block. H. Seedlings after both agar blocks have been applied and the plate has been sealed with Parafilm. I. Marks on the back of the plate indicating the positions of the primary root tips at the time of agar sandwich assembly. - Repeat the above steps with the remainder of the germinated seedlings. Ensure that all possible media combinations are used.
Note: Possible combinations are (block side/plate side): control/control, treatment/control, control/treatment, and treatment/treatment. - Incubate plates vertically in a growth chamber at 29 °C, 16 h light/8 h dark cycle for 4-5 days, or until lateral roots have emerged along the lengths of the primary roots within the sandwiches.
- If any kernels have not germinated, or their primary roots are too short to put in agar sandwiches, check them again after one day.
- Leave the kernels on the germination paper plates. Cover them back up with a paper towel, reseal the plates with Parafilm, and incubate them in a growth chamber.
- Assemble more agar sandwiches if possible on the following day.
Note: Maize kernels sterilized with the above method that do not germinate after 3 days are typically non-viable. Germination rates after 3 days can range from 70-100% given the quality of the seeds used.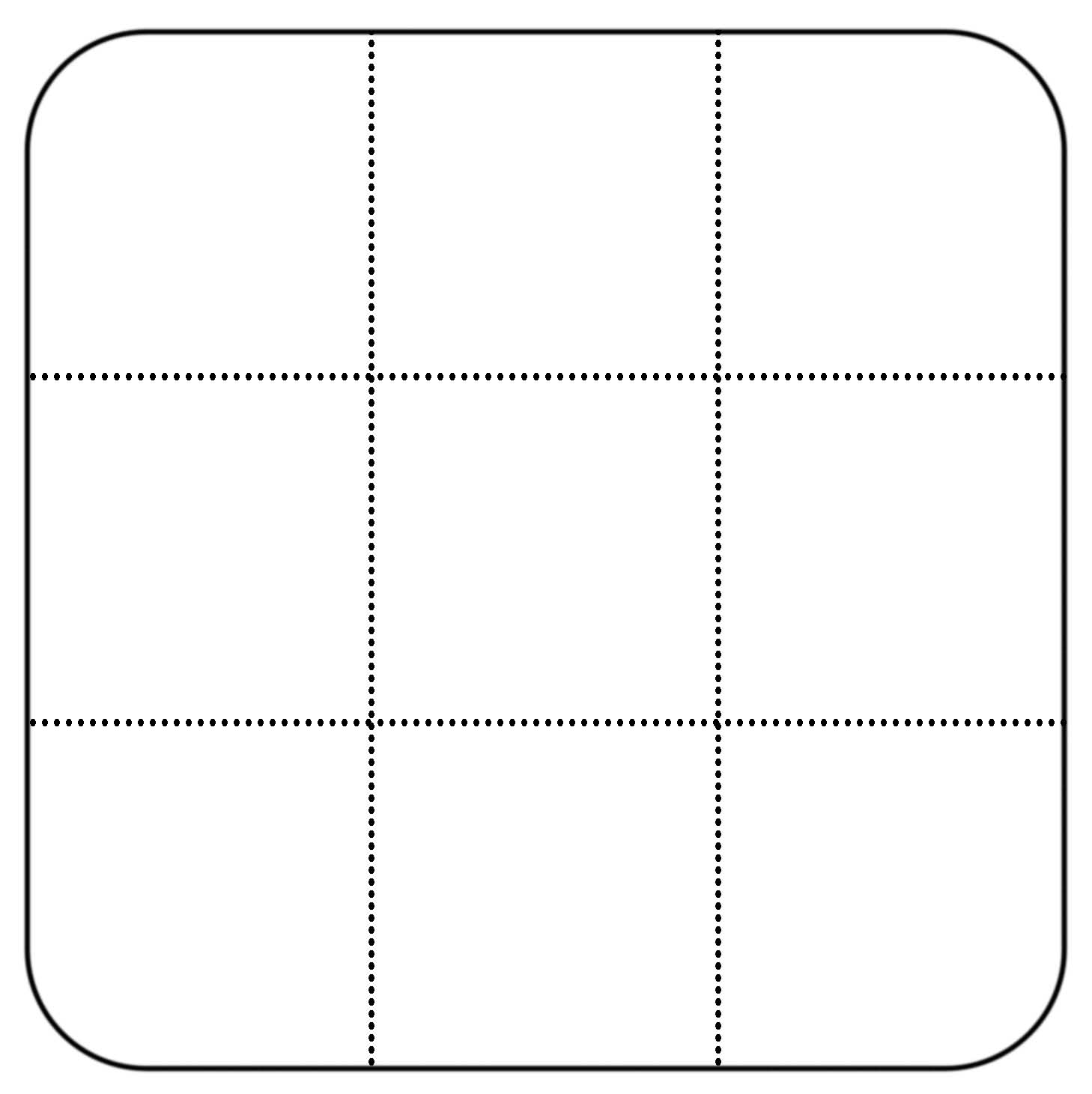
Figure 3. Printable template for cutting agar blocks from Petri dishes. This figure can be printed on an 8.5 x 11-inch sheet of paper and placed underneath Petri dishes to serve as a guide while cutting agar blocks (see step C7).
- Leave the kernels on the germination paper plates. Cover them back up with a paper towel, reseal the plates with Parafilm, and incubate them in a growth chamber.
- Conduct all steps in a laminar flow hood. Spray down all working surfaces and equipment with 70% ethanol before placing into the hood. Use forceps to manipulate materials such as agar, filter paper, silicone rubber, etc.
- Quantification of lateral root patterning from seedlings grown in an agar sandwich
- Remove the Parafilm from the Petri plate and open (Figure 4A).
- Using a permanent marker, mark on the primary roots where they first grew past the agar blocks (Figure 4B, white arrow).
- Carefully remove the agar blocks and filter paper using forceps and a razor blade.
- Use a permanent marker to mark the primary roots where the root tips were at the time of sandwich assembly (denoted on the backs of the plates) (Figure 4B, black arrow). Measure and record the distance between this mark and the mark where the root grew past the agar block with a ruler.
- Count the number of lateral roots that emerged on the side of the primary root that was facing the agar block (Figure 4C and 4D). Only count lateral roots that emerged between the two marks made in steps D2 and D4 (Figure 4C, white and black arrows).
Note: If visualization of lateral roots using the naked eye is difficult, a low-magnification microscope can be used to assist in quantification. - Carefully lift up the primary root and/or rotate the seedling and count the number of lateral roots that emerged on the side exposed to the media in the plate.
Note: Some varieties of maize will accumulate anthocyanin pigmentation in air-exposed regions of the root (Figure 4C and 4D). This can serve as a useful boundary between the side of the root exposed to the block and the side exposed to the media in the plate.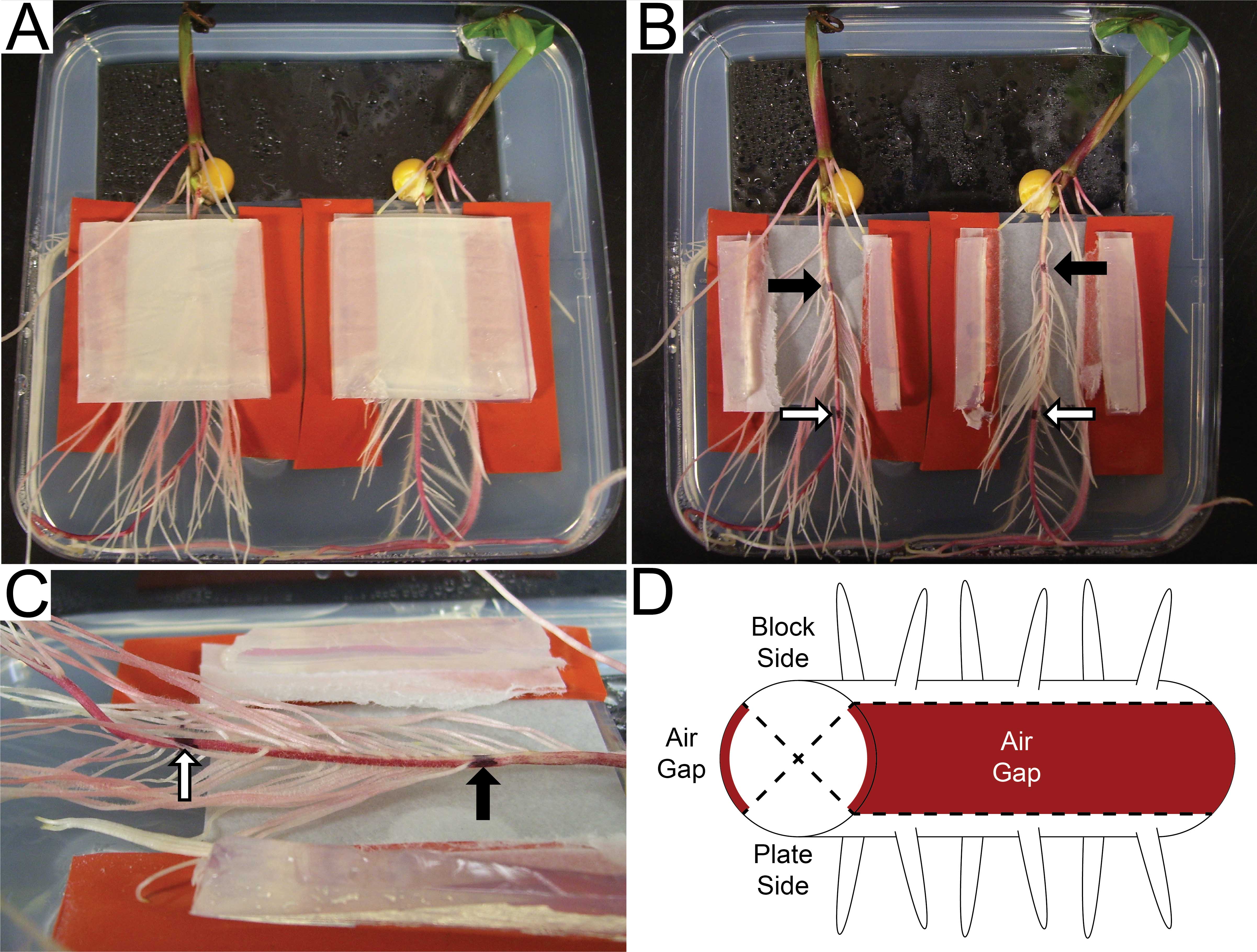
Figure 4. Disassembly of agar sandwiches and plant phenotyping. A. Plants in agar sandwiches five days after sandwich assembly (plate lid removed). B. Primary roots after removal of the agar blocks and overlying filter paper squares. Marks have been placed on the primary roots corresponding to the position of the root tip at the time of sandwich assembly (black arrows) and the point at which the root grew past the agar block (white arrows). C. Side view of the primary root. Lateral root quantification is done between the two marks on the primary root (denoted with arrows as in B). Two lateral root-forming zones are separated by a stripe of red anthocyanin pigmentation that developed in response to air exposure in the gap between the two sides of the sandwich. D. Diagrammatic representation of the different zones of the primary root. Lateral roots appear on the faces of the root exposed to agar media (Block Side and Plate Side), but not in the air-exposed regions (Air Gaps) which are marked with anthocyanin. - Count the number of lateral roots that emerged on the sides of the root exposed to the air gap that was between the agar block and the media in the plate. Only count lateral roots between the two ink marks as described in step D5.
Note: This count is usually close to 0 for most varieties of maize due to lateral root hydropatterning (Bao et al., 2014). - Divide the number of lateral roots on each side of the root by the distance between the two marks (recorded in step D4) to calculate the lateral root density in units of lateral roots/cm.
- Average the lateral root density on each side of the root within each sandwich condition.
- Perform statistical analysis on lateral root density values. The steps to accomplish this are described in brief below. An example script written in R (R Core Team, 2015) has been provided with this protocol (Supplementary files 1 and 2).
- Reformat the data table for use in a linear model (Figure 5)
- The original data table contains separate columns for lateral root density on each side of the root. Convert this to a format in which density and side of the root are in separate columns. This can be accomplished using the “gather()” function in the “tidyr” package (Wickham, 2015). Once this is done, each row of the data table corresponds to a particular lateral root density value.
- Divide the condition column into two columns for the treatment on either side of the sandwich.
- Label the conditions on either side of the sandwich as either “cis” (on the same side of the root as the density value for the current row) or “trans” (on side of the root opposite that of the density value for the current row).
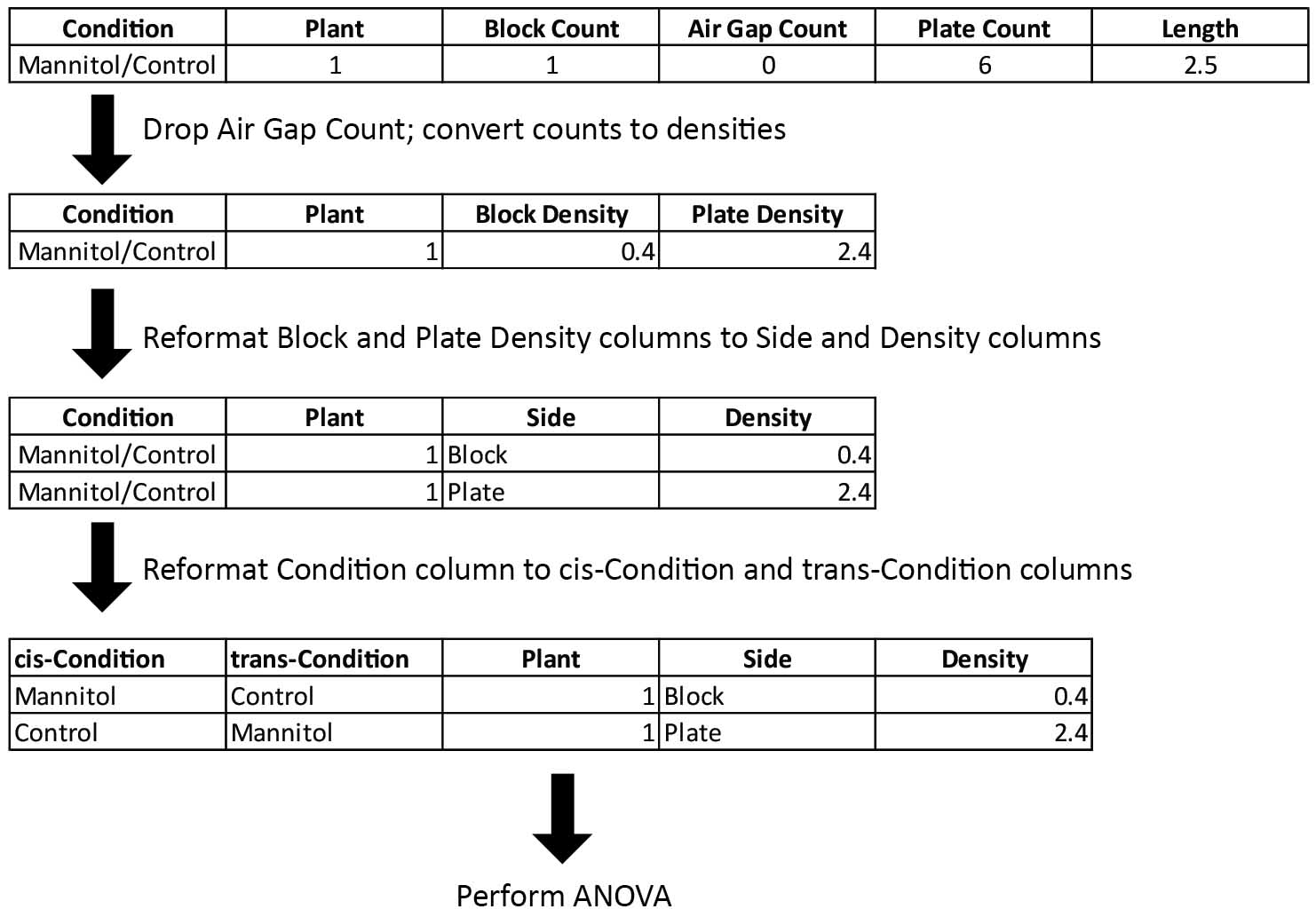
Figure 5. Pre-processing of lateral root quantification data for ANOVA. One plant from one condition is shown as an example. Air Gap Count is dropped from the table since all counts were 0 in each condition, and lateral root densities were calculated by dividing counts by the length of the quantified region. Separate columns for the side of the root and density on that side were then created. Lastly, the condition column was reformatted as cis- and trans-condition categories. Note that the condition notation for the “Block” row is now the inverse of the notation for the “Plate” row. - Perform an ANOVA to assess the effects of side of the root (block vs plate), cis-condition (control vs treatment), trans-condition (control vs treatment), and all possible interaction terms on lateral root density.
- The full linear model is written as: lateral root density ~side + cis-condition + trans-condition + side:cis-condition + side:trans-condition + cis-condition:trans-condition + side:cis-condition:trans-condition + residual error.
- Since each plant in the data set is measured twice (once on each side of the sandwich), an error term to account for within-plant variation must be included. This can be done using the “nlme” package (Pinheiro et al., 2015).
- The ANOVA test will assign P-values for statistical significance for each term in the model. A P-value less than 0.05 can be accepted as statistically significant. The interpretation of significance of each term in the model is explained below.
- Analyze the output from the ANOVA test
- A significant effect of the side of the root (block vs plate), either alone or as an interaction with other variables, indicates that the two sides of the sandwich differ from one another. This may be an indicator of a technical artifact resulting from improper sandwich assembly.
- A significant effect of the cis-condition term indicates that the treatment has a local effect on lateral root development.
- A significant effect of the trans-condition term indicates that the treatment has a non-local effect on lateral root development. This is one indicator that treatment applied to the roots is having systemic effects on lateral root patterning.
- A significant cis-condition:trans-condition interaction term indicates that the response on one side of the root depends upon the conditions experienced on the other side. This is another indicator that the two sides are not responding autonomously to the local treatment.
- If any significant terms are observed from the ANOVA test, perform pairwise comparisons between conditions of interest. This can be done using the “multcomp” package (Hothorn et al., 2008).
- Reformat the data table for use in a linear model (Figure 5)
- Remove the Parafilm from the Petri plate and open (Figure 4A).
Data analysis
See Figure 6A for an example graph of average lateral root densities in different agar sandwich conditions. Table 1 contains the output from an ANOVA conducted in R. Figure 6B summarizes results of post-hoc testing after observation of a significant cis-condition:trans-condition interaction term in the ANOVA.
Table 1. Output from analysis of variance in R. The cis-condition had a significant effect on lateral root density (P < 0.05), which supports the hypothesis that the treatment has a local effect on development. The trans-condition and the cis-condition:trans-condition interaction term were also significant, indicating that the treatment also had a systemic effect on development. All terms which contain “side” as a variable are not significant (P > 0.05), indicating that the effects of the treatment did not depend on the method of application in the agar sandwich (block or plate agar media). numDF and denDF denote numerator and denominator degrees of freedom, respectively.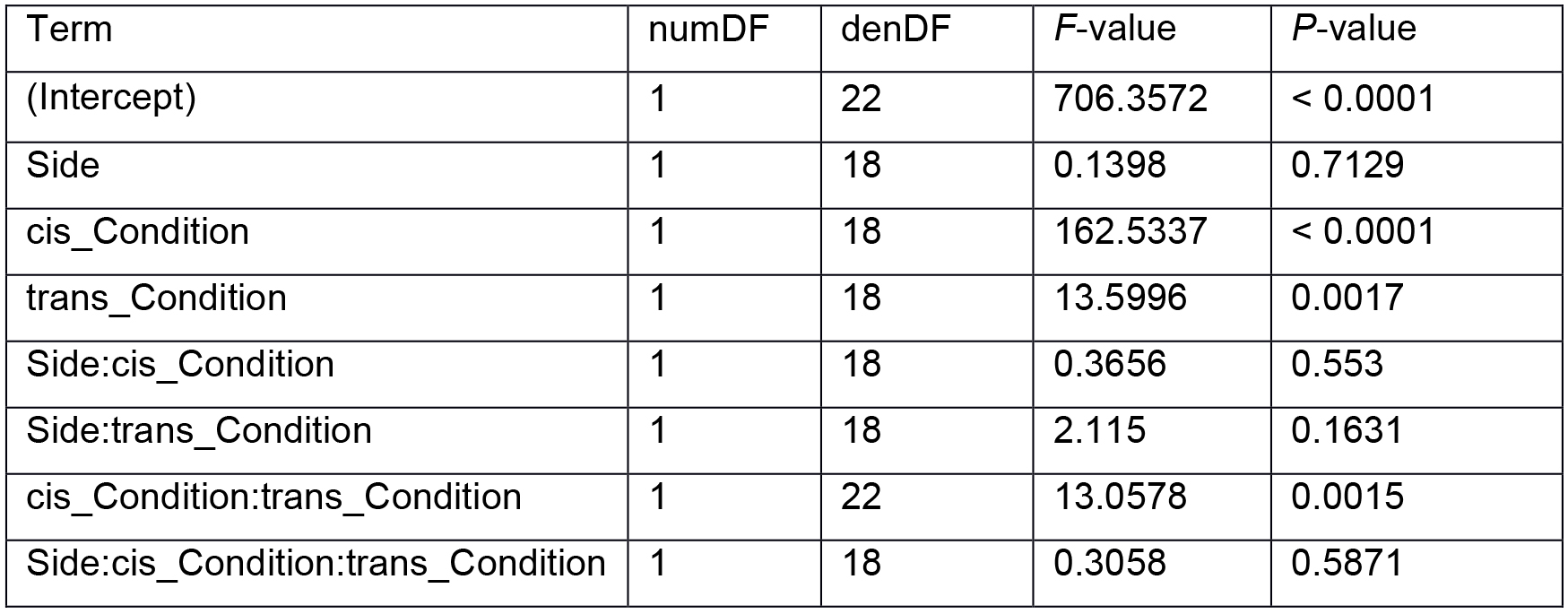
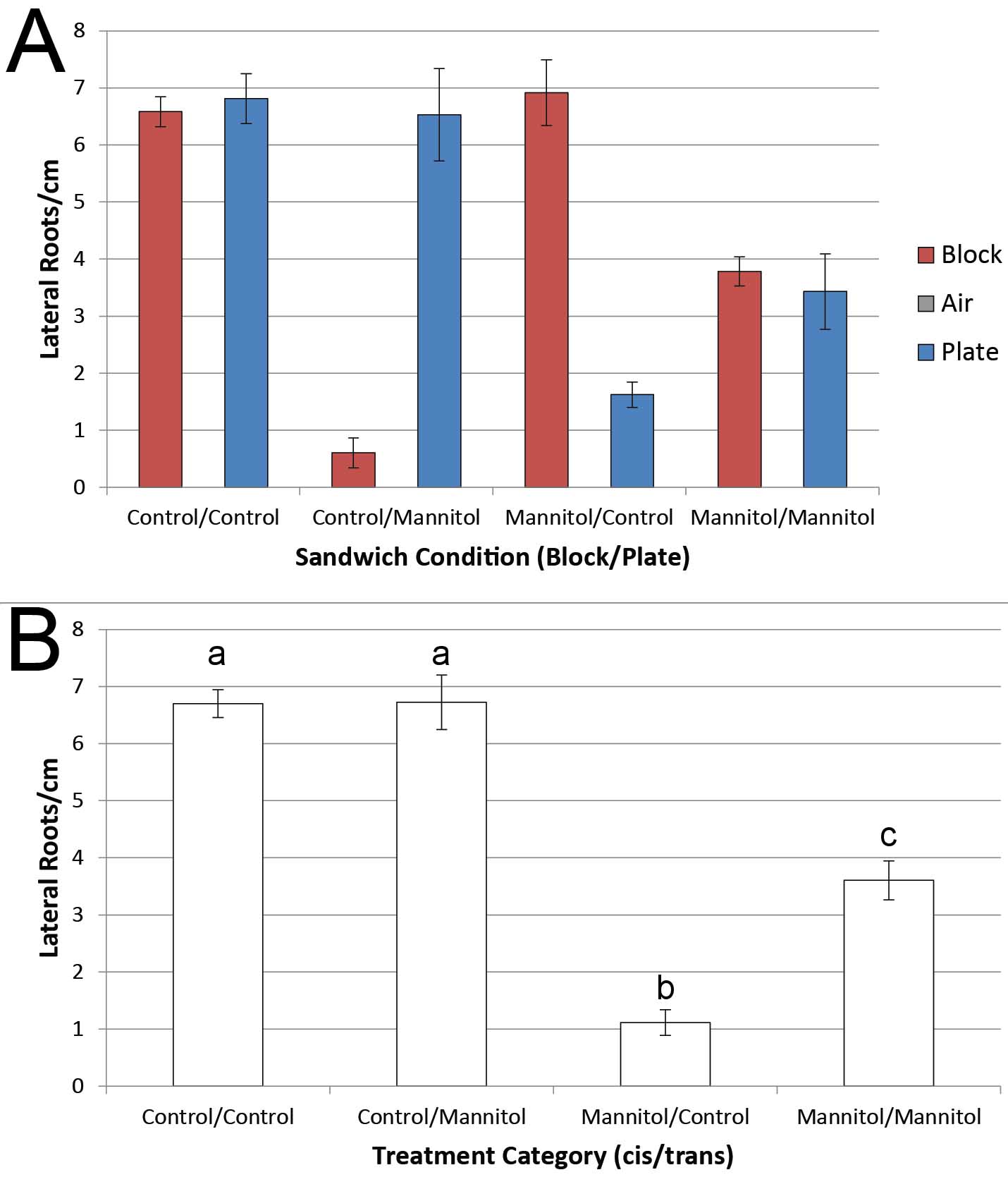
Figure 6. Representative data and statistical analysis. A. Maize seedlings were treated with either control or mannitol agar media in 4 agar sandwich conditions. Average lateral root densities on each side of the root are shown. The Air Gap density was 0 in each condition. Error bars = standard error, n = 6 plants per condition. B. Summary of data from post-hoc testing. Since there was a significant effect of the cis-condition:trans-condition term on lateral root density and no significant effect of the side of the sandwich (block vs plate) (see Table 1), all pairwise comparisons between the different treatment categories were performed using the "multcomp" package in R. Similar groups are denoted with the same letter (P < 0.05 for significant difference). The response to control media was independent of the treatment on the other side of the root, indicating that the root responded locally to this condition. Contrastingly, the response to mannitol differs depending on whether the other side of the root is treated with mannitol or control media, indicating that mannitol can have systemic effects on development. Error bars = standard error, n = 12 observations per category.
Notes
- The agar sandwich procedure was adapted from a procedure used with rice seedlings (Karahara et al., 2012).
- We found that combining heat treatment along with incubation in bleach to be the most effective method of maize kernel sterilization. The heat treatment protocol was adapted from Daniels (1983). The sterilization and seed germination conditions described here have been optimized for maize kernels. Studies involving other plant species may require modifications to these steps to ensure adequate sterility and germination rates (e.g., omission of the heat treatment, addition of an incubation in ethanol, and/or use of different types of germination media to support growth of early seedlings).
- This method is broadly applicable to looking at gradients of essentially any chemical agent that can be applied in gel media. While we have primarily focused on differences in water availability by applying osmolytes on the treatment side (e.g., mannitol or polyethylene glycol), varying concentrations of nutrients such as nitrate and phosphate could also be investigated.
- Applying this technique to other plant species is mainly limited by the diameter of the primary root. The root diameter must be greater than or equal to the thickness of the silicone rubber used to separate the two agar media in the sandwich. If the root is thinner than gap between the two sides of the sandwich, then adequate contact with both media conditions will not occur. Narrower materials could be used as spacers between the two agar media. However, this increases the likelihood of a film of water forming between the two agar surfaces, which would gradually dissipate any chemical gradients across the sandwich.
- If the two media on either side of the sandwich come into direct contact with one another, any differences in concentrations of water-soluble chemicals will equilibrate over time. In order to control for this possibility when examining the effects of different water potentials, we measured the water potential of the media on either side of the sandwich using a vapor pressure osmometer to demonstrate that it was stable during the experiment (Bao et al., 2014). Studies of other chemical gradients may require alternative methods of quality control.
- In our hands, the side of the root exposed to the applied agar block occasionally has fewer lateral roots than the other side, even when the media composition is the same. We attribute this to inadequate contact of the block with the root, as evidenced by increased anthocyanin accumulation on the block side (an indicator of air exposure in maize primary roots). Significant effects of the “side” term in an ANOVA test can also help to identify when this occurs. This can be compensated by increasing the sample size in each sandwich condition, and by taking greater care during assembly of the agar sandwiches.
- The use of filter paper to separate the agar from the root is done to prevent the root tip from growing into either side of the sandwich. The filter paper can be omitted, but the likelihood of the root penetrating into the agar media increases. Additional caution should be exercised to ensure that the two types of agar on either side of the sandwich do not contact one another (see Note 5).
Recipes
- Control agar media
- ½x MS salts + 0.5 g/L MES hydrate
- Adjust pH to 5.7 using potassium hydroxide solution.
- Add 2% (w/v) agar and autoclave for 30 min at 121 °C on wet cycle.
- Pour media into plastic plates in laminar flow hood after cooling, 65 ml/plate. Do not discard the plastic sleeve used for shipping the plates. Volumes can be measured using a sterile 50 ml conical tube.
- After media solidifies, wrap plates in plastic sleeves provided with the plates and store inverted at 4 °C.
- ½x MS salts + 0.5 g/L MES hydrate
- Treatment agar media
- ½x MS salts + 0.5 g/L MES hydrate + 220 mM mannitol
- Adjust pH to 5.7 using potassium hydroxide solution.
- Add 2% (w/v) agar and autoclave for 30 min at 121 °C.
- Pour media into plastic plates and store as above for control agar media.
- ½x MS salts + 0.5 g/L MES hydrate + 220 mM mannitol
- Maize kernel sterilization solution
20% (v/v) bleach + 0.01% (v/v) Tween 20 in distilled water
Prepare fresh immediately before each sterilization. 15 ml is sufficient for up to 30 kernels.
Acknowledgments
This material is based upon work supported by the National Science Foundation Graduate Research Fellowship under Grant No. DGE-1147470. Any opinion, findings, and conclusions or recommendations expressed in this material are those of the authors(s) and do not necessarily reflect the views of the National Science Foundation.
References
- Bao, Y., Aggarwal, P., Robbins, N. E., Sturrock, C. J., Thompson, M. C., Tan, H. Q., Tham, C., Duan, L. N., Rodriguez, P. L., Vernoux, T., Mooney, S. J., Bennett, M. J. and Dinneny, J. R. (2014). Plant roots use a patterning mechanism to position lateral root branches toward available water. Proc Natl Acad Sci U S A 111(25): 9319-9324.
- Daniels, B. A. (1983). Elimination of Fusarium moniliforme from corn seed. Plant Disease 67(6): 609-611.
- Hothorn, T., Bretz, F. and Westfall, P. (2008). Simultaneous inference in general parametric models. Biom J 50(3): 346-363.
- Karahara, I., Umemura, K., Soga, Y., Akai, Y., Bando, T., Ito, Y., Tamaoki, D., Uesugi, K., Abe, J., Yamauchi, D. and Mineyuki, Y. (2012). Demonstration of osmotically dependent promotion of aerenchyma formation at different levels in the primary roots of rice using a 'sandwich' method and X-ray computed tomography. Ann Bot 110(2): 503-509.
- Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., Heisterkamp, S. Willigen, B. V. and R-Core (2016). nlme: linear and nonlinear mixed effects models. R package version 3.1-128.
- R Development Core Team. (2015). R: A language and environment for statistical computing. R Foundation for Statistical Computing.
- Wickham, H. (2016). tidyr: easily tidy data with spread() and gather() functions. R package version 0.6.0.
Article Information
Copyright
© 2016 The Authors; exclusive licensee Bio-protocol LLC.
How to cite
Robbins II, N. E. and Dinneny, J. (2016). A Method to Analyze Local and Systemic Effects of Environmental Stimuli on Root Development in Plants. Bio-protocol 6(17): e1923. DOI: 10.21769/BioProtoc.1923.
Category
Plant Science > Plant physiology > Abiotic stress
Plant Science > Plant physiology > Plant growth
Do you have any questions about this protocol?
Post your question to gather feedback from the community. We will also invite the authors of this article to respond.